Cosa sono le proteine? Corso: Studio degli elementi della composizione chimica dei prodotti alimentari utilizzando l'esempio delle proteine Elementi chimici inclusi nella composizione delle proteine.
Le proprietà di base delle proteine dipendono dalla loro struttura chimica. Le proteine sono composti ad alto peso molecolare le cui molecole sono costituite da residui di alfa aminoacidi, cioè amminoacidi in cui il gruppo amminico primario e il gruppo carbossilico sono collegati allo stesso atomo di carbonio (il primo atomo di carbonio che conta dal gruppo carbonile).
Dalle proteine mediante idrolisi si isolano 19-32 tipi di alfa aminoacidi, ma solitamente si ottengono 20 alfa aminoacidi (questi sono i cosiddetti proteogenico aminoacidi). La loro formula generale:
parte comune a tutti gli aminoacidi
R è un radicale, cioè un raggruppamento di atomi in una molecola di amminoacido associato a un atomo di carbonio alfa e che non partecipa alla formazione della struttura portante di una catena polipeptidica.
Tra i prodotti dell'idrolisi di molte proteine, sono stati trovati prolina e idrossiprolina, che contengono un gruppo amminico =NH, e non un gruppo amminico H 2 N- e sono in realtà imminoacidi, non aminoacidi.
Gli amminoacidi sono sostanze cristalline incolori che fondono e si decompongono ad alte temperature (superiori a 250°C). Facilmente solubile, per la maggior parte, in acqua e insolubile in etere e altri solventi organici.
Gli amminoacidi contengono contemporaneamente due gruppi capaci di ionizzazione: un gruppo carbossilico, che ha proprietà acide, e un gruppo amminico, che ha proprietà basiche, cioè gli amminoacidi sono elettroliti anfoteri.
Nelle soluzioni fortemente acide, gli amminoacidi sono presenti sotto forma di ioni caricati positivamente e in soluzioni alcaline - sotto forma di ioni negativi.
A seconda del valore del pH dell'ambiente, qualsiasi amminoacido può avere una carica positiva o negativa.
Il valore di pH al quale le particelle di amminoacidi sono elettricamente neutre è designato come il loro punto isoelettrico.
Tutti gli amminoacidi derivati da proteine, ad eccezione della glicina, sono otticamente attivi poiché contengono un atomo di carbonio asimmetrico in posizione alfa.
Dei 17 aminoacidi proteici otticamente attivi, 7 sono caratterizzati da rotazione destra /+/ e 10 da rotazione sinistra /-/ del piano del fascio polarizzato, ma tutti appartengono alla serie L.
Gli amminoacidi della serie D sono stati trovati in alcuni composti naturali e oggetti biologici (ad esempio, nei batteri e negli antibiotici gramicidina e actinomicina). Il significato fisiologico degli amminoacidi D e L è diverso. Gli amminoacidi della serie D, di regola, non vengono affatto assorbiti dagli animali e dalle piante, oppure sono scarsamente assorbiti, poiché i sistemi enzimatici degli animali e delle piante sono specificamente adattati agli amminoacidi L. È interessante notare che gli isomeri ottici possono essere distinti in base al gusto: gli amminoacidi della serie L sono amari o insapori e gli amminoacidi della serie D sono dolci.
Tutti i gruppi di amminoacidi sono caratterizzati da reazioni alle quali prendono parte gruppi amminici o gruppi carbossilici, o entrambi. Inoltre, i radicali aminoacidici sono capaci di una varietà di interazioni. I radicali degli aminoacidi reagiscono:
Formazione di sale;
Reazioni redox;
Reazioni di acilazione;
Esterificazione;
Ammidazione;
Fosforilazione.
Queste reazioni, che portano alla formazione di prodotti colorati, sono ampiamente utilizzate per l'identificazione e la determinazione semiquantitativa di singoli aminoacidi e proteine, ad esempio la reazione della xantoproteina (ammidazione), la reazione di Millon (formazione del sale), la reazione del biureto (sale formazione), reazione della ninidrina (ossidazione), ecc.
Anche le proprietà fisiche dei radicali aminoacidici sono molto diverse. Ciò riguarda, prima di tutto, il loro volume e il loro costo. La diversità dei radicali aminoacidici nella natura chimica e nelle proprietà fisiche determina le caratteristiche multifunzionali e specifiche delle proteine che formano.
La classificazione degli amminoacidi presenti nelle proteine può essere effettuata secondo vari criteri: la struttura dello scheletro di carbonio, il contenuto dei gruppi -COOH e H 2 N, ecc. La classificazione più razionale si basa sulle differenze nella polarità degli amminoacidi radicali acidi a pH 7, cioè ad un valore di pH corrispondente alle condizioni intracellulari. In base a ciò, gli amminoacidi che compongono le proteine possono essere suddivisi in quattro classi:
Amminoacidi con radicali apolari;
Amminoacidi con radicali polari vuoti;
Amminoacidi con radicali polari caricati negativamente;
Amminoacidi con radicali polari caricati positivamente
Diamo un'occhiata alla struttura di questi aminoacidi.
Amminoacidi con gruppi R non polari (radicali)
Questa classe comprende quattro amminoacidi alifatici (alanina, valina, isoleucina, leucina), due amminoacidi aromatici (fenilalanina, triptofano), un amminoacido contenente zolfo (metionina) e un amminoacido (prolina). Una proprietà comune di questi amminoacidi è la loro minore solubilità in acqua rispetto agli amminoacidi polari. La loro struttura è la seguente:
Alanina (acido α-amminopropionico)
Valina (acido α-amminoisovalerico)
Leucina (acido α-amminoisocaproico)
Isoleucina (acido α-ammino-β-metilvalerico)
Fenilalanina (acido α-ammino-β-fenilpropionico)
Triptofano (acido α-ammino-β-indolepropionico)
Metionina (acido α-ammino-γ-metil-tiobutirico)
Prolina (acido pirrolidina-α-carbossilico)
2. Amminoacidi con gruppi R polari non carichi (radicali)
Questa classe comprende un amminoacido alifatico - glicina (glicocolo), due amminoacidi idrossilici - serina e treonina, un amminoacido contenente zolfo - cisteina, un amminoacido aromatico - tirosina e due ammidi - asparagina e glutammina.
Questi amminoacidi sono più solubili in acqua rispetto agli amminoacidi con gruppi R non polari perché i loro gruppi polari possono formare legami idrogeno con le molecole d'acqua. La loro struttura è la seguente:
Glicina o glicocole (acido α-amminoacetico)
Serina (acido α-ammino-β-idrossipropionico)
Treonina (acido α-ammino-β-idrossibutirrico)
Cisteina (acido α-ammino-β-tiopropionico)
Tirosina (acido α-ammino-β-paraidrossifenilpropionico)
Asparagina
Aminoacidi - componenti strutturali proteine.Proteine, O proteine(Greco protos - primario) sono eteropolimeri biologici, i cui monomeri sono amminoacidi.
Gli amminoacidi sono composti organici a basso peso molecolare contenenti gruppi carbossilici (-COOH) e amminici (-NH 2) legati allo stesso atomo di carbonio. All'atomo di carbonio è attaccata una catena laterale, un radicale che conferisce a ciascun amminoacido determinate proprietà. La formula generale degli aminoacidi è:

La maggior parte degli amminoacidi ha un gruppo carbossilico e un gruppo amminico; vengono chiamati questi amminoacidi neutro. Ci sono però anche amminoacidi basici- con più di un gruppo amminico, nonché amminoacidi acidi- con più di un gruppo carbossilico.
Sono noti circa 200 amminoacidi presenti negli organismi viventi, ma solo 20 di essi si trovano nelle proteine. Questi sono i cosiddetti di base, O formazione di proteine(proteinogenico), amminoacidi.
A seconda del tipo di radicale, gli amminoacidi basici si dividono in tre gruppi: 1) non polari (alanina, metionina, valina, prolina, leucina, isoleucina, triptofano, fenilalanina); 2) polari scariche (asparagina, glutammina, serie, glicina, tirosina, treonina, cisteina); 3) carica polare (arginina, istidina, lisina - positivi; acidi aspartico e glutammico - negativi).
Le catene laterali degli amminoacidi (radicali) possono essere idrofobiche o idrofile, il che conferisce alle proteine le proprietà corrispondenti che si manifestano nella formazione di strutture proteiche secondarie, terziarie e quaternarie.
Nelle piante Tutto gli amminoacidi essenziali sono sintetizzati dai prodotti primari della fotosintesi. Gli esseri umani e gli animali non sono in grado di sintetizzare un certo numero di aminoacidi proteogenici e devono riceverli in forma finita insieme al cibo. Questi aminoacidi sono chiamati insostituibile. A tra questi ricordiamo lisina, valina, leucina, isoleucina, treonina, fenilalanina, triptofano, metionina; così come arginina e istidina - essenziali per i bambini,
In soluzione gli amminoacidi possono agire sia come acidi che come basi, cioè sono composti anfoteri. Il gruppo carbossilico -COOH è in grado di donare un protone, funzionando come un acido, e il gruppo amminico - NH2 - può accettare un protone, esibendo così le proprietà di una base.
Peptidi. Il gruppo amminico di un amminoacido è in grado di reagire con il gruppo carbossilico di un altro amminoacido.
La molecola risultante è un dipeptide e il legame -CO-NH- è chiamato legame peptidico:

Ad un'estremità della molecola del dipeptide c'è un gruppo amminico libero e all'altra c'è un gruppo carbossilico libero. Grazie a ciò, il dipeptide può legare a sé altri amminoacidi, formando oligopeptidi. Se molti amminoacidi vengono combinati in questo modo (più di dieci), risulta polipeptide.
I peptidi svolgono un ruolo importante nel corpo. Molti oligo e polipeptidi sono ormoni, antibiotici e tossine.
Gli oligopeptidi includono l’ossitocina, la vasopressina, la tireotropina, nonché la bradichinina (peptide del dolore) e alcuni oppiacei (“farmaci naturali” umani) che agiscono come antidolorifici. L'assunzione di farmaci distrugge il sistema oppioide del corpo, quindi un tossicodipendente senza una dose di farmaci sperimenta un forte dolore - "astinenza", che normalmente viene alleviato dagli oppiacei. Anche alcuni antibiotici (ad esempio la gramicidina S) appartengono agli oligopeptidi.
Molti ormoni (insulina, ormone adrenocorticotropo, ecc.), Antibiotici (ad esempio gramicidina A), tossine (ad esempio tossina difterica) sono polipeptidi.
Le proteine sono polipeptidi, la cui molecola contiene da cinquanta a diverse migliaia di amminoacidi con un peso molecolare relativo superiore a 10.000.
Struttura delle proteine. Ogni proteina in un determinato ambiente è caratterizzata da una speciale struttura spaziale. Quando si caratterizza la struttura spaziale (tridimensionale), si distinguono quattro livelli di organizzazione delle molecole proteiche (Fig. 1.1).
|
lie-glu-tre-ala-ala-ala-liz-fen-glu-arg-gln-gis-met-asp-ser- |
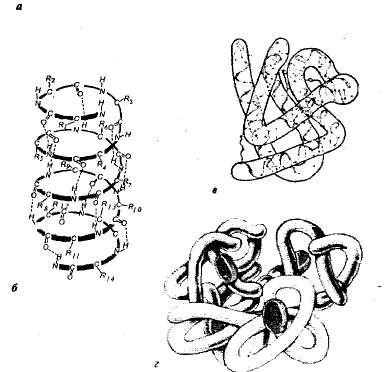
Riso. 1.1. Livelli di organizzazione strutturale delle proteine: a — struttura primaria - sequenza aminoacidica della proteina ribonucleasi (124 unità aminoacidiche); B — struttura secondaria — la catena polipeptidica è attorcigliata a forma di spirale; V— struttura terziaria della proteina mioglobina; G — struttura quaternaria dell'emoglobina.
Struttura primaria— la sequenza degli amminoacidi in una catena polipeptidica. Questa struttura è specifica per ciascuna proteina ed è determinata dall'informazione genetica, cioè dipende dalla sequenza di nucleotidi nella sezione della molecola di DNA che codifica per quella proteina. Tutte le proprietà e le funzioni delle proteine dipendono dalla struttura primaria. La sostituzione di un singolo amminoacido nella composizione delle molecole proteiche o l'interruzione della loro disposizione comporta solitamente un cambiamento nella funzione della proteina.
Considerando che le proteine contengono 20 tipi di aminoacidi, il numero di opzioni per le loro combinazioni nella catena polipeptidica è davvero illimitato, il che fornisce un numero enorme di tipi di proteine nelle cellule viventi. Ad esempio, nel corpo umano sono state trovate più di 10mila proteine diverse, tutte costituite dagli stessi 20 amminoacidi fondamentali.
Nelle cellule viventi, le molecole proteiche o le singole sezioni di esse non sono una catena allungata, ma sono attorcigliate in una spirale, che ricorda una molla estesa (questa è la cosiddetta a-elica), o piegate in uno strato piegato (p- strato). Tali a-eliche e p-fogli sono secondari struttura. Si verifica come risultato della formazione di legami idrogeno all'interno di una catena polipeptidica (configurazione elicoidale) o tra due catene polipeptidiche (strati ripiegati).
La proteina cheratina ha una configurazione completamente aelicoidale. È la proteina strutturale di capelli, unghie, artigli, becchi, piume e corna; fa parte dello strato esterno della pelle dei vertebrati.
Nella maggior parte delle proteine, le sezioni elicoidali e non elicoidali della catena polipeptidica si ripiegano in una formazione sferica tridimensionale - un globulo (caratteristico delle proteine globulari). Un globulo di una certa configurazione lo è struttura terziaria scoiattolo. Questa struttura è stabilizzata da legami ionici, idrogeno, disolfuro covalenti (formati tra atomi di zolfo che fanno parte di cisteina, cistina e megionina), nonché da interazioni idrofobiche. Le più importanti nell'emergere della struttura terziaria sono le interazioni idrofobiche; In questo caso, la proteina si piega in modo tale che le sue catene laterali idrofobe siano nascoste all'interno della molecola, cioè siano protette dal contatto con l'acqua, e le catene laterali idrofile, al contrario, siano esposte all'esterno.
Molte proteine con una struttura particolarmente complessa sono costituite da diverse catene polipeptidiche (subunità), che si formano struttura quaternaria molecola proteica. Questa struttura si trova, ad esempio, nella proteina globulare dell'emoglobina. La sua molecola è costituita da quattro subunità polipeptidiche separate (protomeri), situate nella struttura terziaria, e da una parte non proteica: l'eme.
Solo in questa struttura l'emoglobina è in grado di svolgere la sua funzione di trasporto.
Sotto l'influenza di vari fattori chimici e fisici (trattamento con alcool, acetone, acidi, alcali, alta temperatura, irradiazione, alta pressione, ecc.), si verifica un cambiamento nelle strutture secondaria, terziaria e quaternaria della proteina a causa della rottura di legami idrogeno e ionici. Viene chiamato il processo di distruzione della struttura nativa (naturale) di una proteina denaturazione. In questo caso si verifica una diminuzione della solubilità proteica, un cambiamento nella forma e nella dimensione delle molecole, una perdita di attività enzimatica, ecc. Il processo di denaturazione può essere completo o parziale. In alcuni casi, il passaggio alle normali condizioni ambientali è accompagnato dal ripristino spontaneo della struttura naturale della proteina. Questo processo si chiama rinaturazione.
Proteine semplici e complesse. In base alla loro composizione chimica, le proteine si dividono in semplici e complesse. Perdonami includere proteine costituite solo da aminoacidi e difficile- proteine contenenti una parte proteica ed una parte non proteica (protesi); un gruppo prostetico può essere formato da ioni metallici, un residuo di acido fosforico, carboidrati, lipidi, ecc. Le proteine semplici sono l'albumina sierica, la fibrina, alcuni enzimi (tripsina), ecc. Le proteine complesse comprendono tutti i proteolipidi e le glicoproteine; le proteine complesse sono, ad esempio, le immunoglobuline (anticorpi), l'emoglobina, la maggior parte degli enzimi, ecc.
Funzioni delle proteine.
- Strutturale. Le proteine fanno parte delle membrane cellulari e della matrice degli organelli cellulari. Le pareti dei vasi sanguigni, della cartilagine, dei tendini, dei capelli, delle unghie e degli artigli negli animali superiori sono costituite principalmente da proteine.
- Catalitico (enzimatico). Le proteine enzimatiche catalizzano tutte le reazioni chimiche nel corpo. Garantiscono la scomposizione dei nutrienti nel tratto digestivo, la fissazione del carbonio durante la fotosintesi, ecc.
- Trasporto. Alcune proteine sono in grado di legare e trasportare varie sostanze. Le albumine del sangue trasportano gli acidi grassi, le globuline trasportano ioni metallici e ormoni e l'emoglobina trasporta ossigeno e anidride carbonica. Le molecole proteiche che compongono la membrana plasmatica prendono parte al trasporto di sostanze nella cellula.
- Protettivo. Viene eseguito dalle immunoglobuline (anticorpi) nel sangue, che forniscono la difesa immunitaria del corpo. Il fibrinogeno e la trombina sono coinvolti nella coagulazione del sangue e prevengono il sanguinamento.
- Contrattile. A causa dello scorrimento delle protofibrille di actina e miosina l'una rispetto all'altra, si verificano la contrazione muscolare e le contrazioni intracellulari non muscolari. Il movimento delle ciglia e dei flagelli è associato allo scorrimento dei microtubuli di natura proteica l'uno rispetto all'altro.
- Normativa. Molti ormoni sono oligopeptidi o peptidi (p. es., insulina, glucagone [antagonista dell'insulina], ormone adrenocorticotropo, ecc.).
- Recettore. Alcune proteine incorporate nella membrana cellulare sono in grado di modificare la loro struttura sotto l'influenza dell'ambiente esterno. In questo modo vengono ricevuti i segnali dall'esterno e le informazioni vengono trasmesse nella cellula. Un esempio potrebbe essere fitocromo-- una proteina fotosensibile che regola la risposta fotoperiodica delle piante, e opsina - componente rodopsina, pigmento presente nelle cellule della retina.
- Energia. Le proteine possono servire come fonte di energia nella cellula (dopo la loro idrolisi). Tipicamente, le proteine vengono utilizzate per il fabbisogno energetico in casi estremi, quando le riserve di carboidrati e grassi sono esaurite.
Enzimi (enzimi). Si tratta di proteine specifiche presenti in tutti gli organismi viventi e che svolgono il ruolo di catalizzatori biologici.
Le reazioni chimiche in una cellula vivente si verificano a una certa temperatura, pressione normale e acidità appropriata dell'ambiente. In tali condizioni, le reazioni di sintesi o di degradazione delle sostanze procederebbero molto lentamente nella cellula se non fossero esposte agli enzimi. Gli enzimi accelerano una reazione senza modificarne il risultato complessivo riducendo energia di attivazione, cioè, quando sono presenti, è necessaria molta meno energia per rendere reattive le molecole che reagiscono, oppure la reazione procede lungo un percorso diverso con una barriera energetica inferiore.
Tutti i processi in un organismo vivente vengono eseguiti direttamente o indirettamente con la partecipazione di enzimi. Ad esempio, sotto la loro influenza, i componenti costitutivi del cibo (proteine, carboidrati, lipidi, ecc.) Vengono scomposti in composti più semplici e da essi vengono quindi sintetizzate nuove macromolecole caratteristiche di questo tipo. Pertanto, i disturbi nella formazione e nell'attività degli enzimi spesso portano alla comparsa di malattie gravi.
Secondo la loro organizzazione spaziale, gli enzimi sono costituiti da diversi sessi e catene peptidiche e di solito hanno una struttura quaternaria. Inoltre, gli enzimi possono includere anche strutture non proteiche. La parte proteica lo è nome apoenzima, e non proteici - cofattore(se si tratta di cationi o anioni di sostanze inorganiche, ad esempio Zn 2- Mn 2+, ecc.) oppure coenzima (coenzima)(se si tratta di una sostanza organica a basso peso molecolare).
Le vitamine sono precursori o componenti di molti coenzimi. Pertanto, l'acido pantotenico è un componente del coenzima A, l'acido nicotinico (vitamina PP) è un precursore del NAD e del NADP, ecc.
La catalisi enzimatica obbedisce alle stesse leggi della catalisi non enzimatica nell'industria chimica, ma a differenza di essa è caratterizzata da insoliti elevato grado di specificità(un enzima catalizza una sola reazione o agisce solo su un tipo di legame). Ciò garantisce una regolazione precisa di tutti i processi vitali (respirazione, digestione, fotosintesi, ecc.) che si verificano nella cellula e nel corpo. Ad esempio, l'enzima ureasi catalizza la scomposizione di una sola sostanza: l'urea (H 2 N-CO-NH 2 + H 2 O -> -» 2NH 3 + CO 2), senza esercitare un effetto catalitico sui composti strutturalmente correlati.
Per comprendere il meccanismo d'azione degli enzimi ad alta specificità, è molto La teoria del centro attivo è importante. Secondo lei, V molecola tutti enzima ce n'è uno uno o più siti in cui avviene la catalisi a causa dello stretto contatto (in molti punti) tra le molecole dell'enzima e una sostanza specifica (substrato). Il centro attivo è un gruppo funzionale (ad esempio, il gruppo OH della serina) o un amminoacido separato. Tipicamente, un effetto catalitico richiede una combinazione di diversi residui di amminoacidi (in media da 3 a 12) situati in un certo ordine. Il centro attivo è formato anche da ioni metallici, vitamine e altri composti non proteici associati all'enzima: coenzimi o cofattori. Inoltre, la forma e la struttura chimica del centro attivo sono tali Con Solo alcuni substrati possono legarsi ad esso a causa della loro corrispondenza ideale (complementarità o complementarità) tra loro. Il ruolo dei residui amminoacidici rimanenti in una grande molecola enzimatica è quello di fornire alla sua molecola la forma globulare appropriata, necessaria per l'efficace funzionamento del centro attivo. Inoltre, attorno a una grande molecola di enzima si forma un forte campo elettrico. In un tale campo diventa possibile che le molecole del substrato si orientino e acquisiscano una forma asimmetrica. Ciò porta ad un indebolimento dei legami chimici e la reazione catalizzata avviene con un dispendio energetico iniziale minore, e quindi ad una velocità molto più elevata. Ad esempio, una molecola dell'enzima catalasi può scomporre in 1 minuto più di 5 milioni di molecole di perossido di idrogeno (H 2 0 2), che si forma durante l'ossidazione di vari composti nel corpo.
In alcuni enzimi, in presenza di un substrato, la configurazione del centro attivo subisce modifiche, cioè l'enzima orienta i suoi gruppi funzionali in modo da garantire la massima attività catalitica.
Nella fase finale della reazione chimica, il complesso enzima-substrato viene separato per formare i prodotti finali e l'enzima libero. Il centro attivo liberato in questo caso può accettare nuove molecole di substrato.
Velocità delle reazioni enzimatiche dipende da molti fattori: natura e concentrazione dell'enzima e del substrato, temperatura, pressione, acidità del mezzo, presenza di inibitori, ecc. Ad esempio, a temperature prossime allo zero, la velocità delle reazioni biochimiche rallenta al minimo . Questa proprietà è ampiamente utilizzata in vari settori dell'economia nazionale, in particolare in agricoltura e medicina. In particolare, conservazione Prima del trapianto di vari organi (reni, cuore, milza, fegato) a un paziente, questi vengono raffreddati per ridurre l'intensità delle reazioni biochimiche e prolungare la vita degli organi. Il congelamento rapido dei prodotti alimentari impedisce la crescita e la riproduzione dei microrganismi (batteri, funghi, ecc.) e inattiva anche i loro enzimi digestivi, in modo che non siano più in grado di causare la decomposizione dei prodotti alimentari.
Fonte : N / A. Lemeza L.V.Kamlyuk N.D. Lisov "Un manuale di biologia per chi entra nelle università"
Come sapete, le proteine sono una componente necessaria e basilare di qualsiasi organismo vivente. Sono responsabili del metabolismo e della conversione dell’energia, che sono indissolubilmente legati a quasi tutti i processi vitali. La stragrande maggioranza dei tessuti e degli organi degli animali e dell'uomo, nonché oltre il 50% di tutti i microrganismi, sono costituiti principalmente da proteine (dal 40% al 50%). Inoltre nel mondo vegetale ce ne sono meno rispetto alla media, mentre nel mondo animale sono di più. Tuttavia, la composizione chimica delle proteine è ancora sconosciuta a molte persone. Ricordiamo ancora una volta cosa c'è dentro questi ad alto peso molecolare
Composizione proteica
Questa sostanza contiene in media circa il 50-55% di carbonio, 15-17% di azoto, 21-23% di ossigeno, 0,3-2,5% di zolfo. Oltre ai componenti principali elencati, a volte le proteine contengono elementi il cui peso specifico è molto insignificante. Prima di tutto, si tratta di fosforo, ferro, iodio, rame e alcuni altri micro e macroelementi. È interessante notare che la concentrazione di azoto è la più costante, mentre il contenuto di altri componenti chiave può variare. Nel descrivere la composizione di una proteina, va notato che si tratta di un polimero irregolare, costituito da residui del quale in una soluzione acquosa a pH neutro nella forma più generale può essere scritto come NH3 + CHRCOO-.
Questi “mattoni” sono collegati tra loro da un legame ammidico tra i gruppi carbossilici e amminici. In totale, in natura sono state identificate circa un migliaio di proteine diverse. Questa classe comprende anticorpi, enzimi, molti ormoni e altre sostanze biologiche attive. Sorprendentemente, con tutta questa diversità, la composizione proteica può includere non più di 30 proteine diverse, di cui le più popolari sono le più apprezzate. Il corpo umano ne contiene solo 22, il resto semplicemente non viene assorbito e viene escreto. Otto aminoacidi di questo gruppo sono considerati essenziali. Questi sono leucina, metionina, isoleucina, lisina, fenilalanina, triptofano, treonina e valina. Il nostro corpo non è in grado di sintetizzarli da solo e quindi ne richiede l'approvvigionamento dall'esterno.

Può facilmente creare da solo il resto (taurina, arginina, glicina, carnitina, asparagina, istidina, cisteina, glutammina, alanina, ornitina, tirosina, prolina, serina, cistina). Pertanto, tali aminoacidi sono classificati come non essenziali. A seconda della presenza del primo gruppo di proteine nella composizione, nonché del grado di assorbimento da parte dell'organismo, le proteine si dividono in complete e incomplete. L'assunzione media giornaliera di questa sostanza per una persona varia da 1 a 2 grammi per chilogrammo di peso. In questo caso, le persone sedentarie dovrebbero attenersi al limite inferiore di questo intervallo e gli atleti dovrebbero attenersi al limite superiore.
Come studiare la composizione proteica

Per studiare queste sostanze viene utilizzato principalmente il metodo dell'idrolisi. La proteina di interesse viene riscaldata con acido cloridrico diluito (6-10 mol/litro) ad una temperatura compresa tra 100°C e 1100°C. Di conseguenza, si scompone in una miscela di amminoacidi, dalla quale sono già isolati i singoli amminoacidi. Attualmente, per la proteina in studio vengono utilizzati carta e cromatografia a scambio ionico. Esistono anche speciali analizzatori automatici che possono facilmente determinare quali amminoacidi si formano a seguito della scomposizione.
Contenuto dell'articolo
PROTEINE (Articolo 1)– una classe di polimeri biologici presenti in ogni organismo vivente. Con la partecipazione delle proteine avvengono i principali processi che assicurano le funzioni vitali del corpo: respirazione, digestione, contrazione muscolare, trasmissione degli impulsi nervosi. Il tessuto osseo, la pelle, i capelli e le formazioni cornee degli esseri viventi sono costituiti da proteine. Per la maggior parte dei mammiferi, la crescita e lo sviluppo del corpo avviene attraverso alimenti contenenti proteine come componente alimentare. Il ruolo delle proteine nel corpo e, di conseguenza, la loro struttura è molto varia.
Composizione proteica.
Tutte le proteine sono polimeri le cui catene sono assemblate da frammenti di amminoacidi. Gli amminoacidi sono composti organici contenenti nella loro composizione (secondo il nome) un gruppo amminico NH 2 e un gruppo acido organico, ad es. gruppo carbossilico, COOH. Della varietà di amminoacidi esistenti (teoricamente il numero di amminoacidi possibili è illimitato), solo quelli che hanno un solo atomo di carbonio tra il gruppo amminico e il gruppo carbossilico partecipano alla formazione delle proteine. In generale, gli amminoacidi coinvolti nella formazione delle proteine possono essere rappresentati dalla formula: H 2 N–CH(R)–COOH. Il gruppo R attaccato all'atomo di carbonio (quello tra i gruppi amminico e carbossilico) determina la differenza tra gli aminoacidi che formano le proteine. Questo gruppo può essere costituito solo da atomi di carbonio e idrogeno, ma più spesso contiene, oltre a C e H, vari gruppi funzionali (capaci di ulteriori trasformazioni), ad esempio HO-, H 2 N-, ecc. Esiste anche un'opzione quando R = H.
Gli organismi degli esseri viventi contengono più di 100 aminoacidi diversi, tuttavia non tutti vengono utilizzati nella costruzione delle proteine, ma solo 20, quelli cosiddetti “fondamentali”. Nella tabella 1 mostra i loro nomi (la maggior parte dei nomi si è formata storicamente), la formula strutturale e la designazione abbreviata ampiamente utilizzata. Tutte le formule strutturali sono disposte nella tabella in modo che il frammento principale dell'amminoacido sia sulla destra.
| Nome | Struttura | Designazione |
| GLICINA | GLI | |
| Alanina | ALA | |
| VALINA | LANCIA | |
| LEUCINA | LEI | |
| ISOLEUCINA | ILE | |
| SERINA | SER | |
| TREONINA | TRE | |
| CISTEINA | CIS | |
| METIONINA | INCONTRATO | |
| LISINA | LIZ | |
| ARGININA | ARG | |
| ACIDO ASPARAGICO | ASN | |
| ASPARAGINA | ASN | |
| ACIDO GLUTAMMICO | GLU | |
| GLUTAMMINA | GLN | |
| FENILALANINA | ASCIUGACAPELLI | |
| TIROSINA | TIR | |
| TRIPTOFANO | TRE | |
| ISTIDINA | GIS | |
| PROLINA | PRO | |
| Nella pratica internazionale, è accettata la designazione abbreviata degli amminoacidi elencati utilizzando abbreviazioni latine di tre lettere o di una lettera, ad esempio glicina - Gly o G, alanina - Ala o A. | ||
Tra questi venti amminoacidi (Tabella 1), solo la prolina contiene un gruppo NH accanto al gruppo carbossilico COOH (invece di NH 2), poiché fa parte del frammento ciclico.
Otto aminoacidi (valina, leucina, isoleucina, treonina, metionina, lisina, fenilalanina e triptofano), disposti nella tabella su sfondo grigio, sono chiamati essenziali, poiché il corpo deve riceverli costantemente dagli alimenti proteici per la normale crescita e sviluppo.
Una molecola proteica si forma come risultato della connessione sequenziale di amminoacidi, mentre il gruppo carbossilico di un acido interagisce con il gruppo amminico di una molecola vicina, determinando la formazione di un legame peptidico –CO–NH– e il rilascio di una molecola d'acqua. Nella fig. La Figura 1 mostra una combinazione sequenziale di alanina, valina e glicina.
Riso. 1 COLLEGAMENTO IN SERIE DEGLI AMINOACIDI durante la formazione di una molecola proteica. Il percorso dal gruppo amminico terminale di H 2 N al gruppo carbossilico terminale di COOH è stato scelto come direzione principale della catena polimerica.
Per descrivere in modo compatto la struttura di una molecola proteica, vengono utilizzate le abbreviazioni degli amminoacidi (Tabella 1, terza colonna) coinvolti nella formazione della catena polimerica. Il frammento della molecola mostrato in Fig. 1 si scrive così: H 2 N-ALA-VAL-GLY-COOH.
Le molecole proteiche contengono da 50 a 1500 residui di aminoacidi (le catene più corte sono chiamate polipeptidi). L'individualità di una proteina è determinata dall'insieme degli amminoacidi che compongono la catena polimerica e, cosa non meno importante, dall'ordine della loro alternanza lungo la catena. Ad esempio, la molecola di insulina è composta da 51 residui di aminoacidi (questa è una delle proteine a catena più corta) ed è costituita da due catene parallele di lunghezza disuguale collegate tra loro. L'ordine di alternanza dei frammenti di amminoacidi è mostrato in Fig. 2.
Riso. 2 MOLECOLA DI INSULINA, costituito da 51 residui di amminoacidi, i frammenti di amminoacidi identici sono contrassegnati con un colore di sfondo corrispondente. I residui di amminoacidi cisteina contenuti nella catena (abbreviato CIS) formano ponti disolfuro –S-S-, che collegano due molecole polimeriche, oppure formano ponti all’interno di una catena.
Le molecole di aminoacidi della cisteina (Tabella 1) contengono gruppi solfidrico reattivi –SH, che interagiscono tra loro, formando ponti disolfuro –S-S-. Il ruolo della cisteina nel mondo delle proteine è speciale; con la sua partecipazione si formano legami incrociati tra le molecole proteiche polimeriche.
La combinazione di amminoacidi in una catena polimerica avviene in un organismo vivente sotto il controllo degli acidi nucleici che forniscono un rigoroso ordine di assemblaggio e regolano la lunghezza fissa della molecola polimerica (; cm. ACIDI NUCLEICI).
Struttura delle proteine.
La composizione della molecola proteica, presentata sotto forma di residui aminoacidici alternati (Fig. 2), è chiamata struttura primaria della proteina. I legami idrogeno si verificano tra i gruppi imminici HN e i gruppi carbonilici CO presenti nella catena polimerica ( cm. LEGAME IDROGENO), di conseguenza, la molecola proteica acquisisce una certa forma spaziale, chiamata struttura secondaria. I tipi più comuni di struttura secondaria delle proteine sono due.
La prima opzione, chiamata α-elica, è realizzata utilizzando legami idrogeno all'interno di una singola molecola polimerica. I parametri geometrici della molecola, determinati dalle lunghezze e dagli angoli di legame, sono tali che la formazione di legami idrogeno è possibile per i gruppi H-N e C=O, tra i quali si trovano due frammenti peptidici H-N-C=O (Fig. 3).
La composizione della catena polipeptidica mostrata in Fig. 3, redatto in forma abbreviata come segue:
H 2 N-ALA VAL-ALA-LEY-ALA-ALA-ALA-ALA-VAL-ALA-ALA-ALA-COOH.
Come risultato dell'effetto restrittivo dei legami idrogeno, la molecola assume la forma di una spirale - la cosiddetta α-elica, è raffigurata come un nastro a spirale curvo che passa attraverso gli atomi che formano la catena polimerica (Fig. 4 )
Riso. 4 MODELLO 3D DI UNA MOLECOLA PROTEICA sotto forma di α-elica. I legami idrogeno sono mostrati con linee tratteggiate verdi. La forma cilindrica dell'elica è visibile con un certo angolo di rotazione (gli atomi di idrogeno non sono mostrati nella figura). La colorazione dei singoli atomi è data in accordo con le norme internazionali, che consigliano il nero per gli atomi di carbonio, il blu per l'azoto, il rosso per l'ossigeno, il giallo per lo zolfo (per gli atomi di idrogeno non rappresentati in figura si consiglia il bianco, in questo caso l'intero struttura raffigurata su uno sfondo scuro).
Un'altra variante della struttura secondaria, chiamata struttura β, si forma anche con la partecipazione di legami idrogeno, la differenza è che i gruppi H-N e C=O di due o più catene polimeriche situate in parallelo interagiscono. Poiché la catena polipeptidica ha una direzione (Fig. 1), le opzioni sono possibili quando la direzione delle catene coincide (struttura β parallela, Fig. 5), oppure sono opposte (struttura β antiparallela, Fig. 6).
Catene polimeriche di varia composizione possono partecipare alla formazione della struttura β, mentre i gruppi organici che compongono la catena polimerica (Ph, CH 2 OH, ecc.) nella maggior parte dei casi svolgono un ruolo secondario nella posizione relativa di H-N e C =O gruppi è decisivo. Poiché i gruppi H-N e C=O sono diretti in direzioni diverse rispetto alla catena polimerica (su e giù nella figura), diventa possibile l'interazione simultanea di tre o più catene.
La composizione della prima catena polipeptidica in Fig. 5:
H2N-LEY-ALA-FEN-GLY-ALA-ALA-COOH
Composizione della seconda e terza catena:
H2N-GLY-ALA-SER-GLY-TRE-ALA-COOH
La composizione delle catene polipeptidiche mostrata in Fig. 6, lo stesso della Fig. 5, la differenza è che la seconda catena ha verso opposto (rispetto alla Fig. 5).
La formazione di una struttura β all'interno di una molecola è possibile quando un frammento di catena in una certa area viene ruotato di 180°, in questo caso due rami di una molecola hanno direzioni opposte, determinando la formazione di una struttura β antiparallela (; Figura 7).
La struttura mostrata in Fig. 7 in un'immagine piatta, mostrata in Fig. 8 sotto forma di modello tridimensionale. Le sezioni della struttura β sono solitamente contrassegnate semplicemente da un nastro ondulato piatto che attraversa gli atomi che formano la catena polimerica.
La struttura di molte proteine alterna strutture ad α-elica e strutture β a nastro, nonché singole catene polipeptidiche. La loro reciproca disposizione e alternanza nella catena polimerica è chiamata struttura terziaria della proteina.
I metodi per rappresentare la struttura delle proteine sono mostrati di seguito utilizzando l'esempio della proteina vegetale crambin. Le formule strutturali delle proteine, che spesso contengono fino a centinaia di frammenti di amminoacidi, sono complesse, scomode e difficili da comprendere, quindi a volte vengono utilizzate formule strutturali semplificate - senza simboli di elementi chimici (Fig. 9, opzione A), ma mantenendo allo stesso tempo il colore dei tratti di valenza secondo le norme internazionali (Fig. 4). In questo caso, la formula non è presentata in un'immagine piatta, ma spaziale, che corrisponde alla struttura reale della molecola. Questo metodo consente, ad esempio, di distinguere i ponti disolfuro (simili a quelli presenti nell'insulina, Fig. 2), i gruppi fenilici nella struttura laterale della catena, ecc. L'immagine delle molecole sotto forma di modelli tridimensionali (sfere collegati da aste) è un po' più chiaro (Fig. 9, opzione B). Tuttavia, entrambi i metodi non consentono di mostrare la struttura terziaria, quindi la biofisica americana Jane Richardson ha proposto di rappresentare le strutture α sotto forma di nastri attorcigliati a spirale (vedi Fig. 4), le strutture β sotto forma di nastri ondulati piatti (Fig. 8) e collegandole singole catene - sotto forma di fasci sottili, ogni tipo di struttura ha il suo colore. Questo metodo per rappresentare la struttura terziaria di una proteina è ora ampiamente utilizzato (Fig. 9, opzione B). A volte, per maggiore informazione, vengono mostrate insieme la struttura terziaria e una formula strutturale semplificata (Fig. 9, opzione D). Ci sono anche modifiche al metodo proposto da Richardson: le eliche α sono rappresentate come cilindri e le strutture β sono rappresentate sotto forma di frecce piatte che indicano la direzione della catena (Fig. 9, opzione E). Un metodo meno comune è quello in cui l'intera molecola è raffigurata sotto forma di una corda, dove le strutture disuguali sono evidenziate con colori diversi e i ponti disolfuro sono mostrati come ponti gialli (Fig. 9, opzione E).
La più conveniente per la percezione è l'opzione B, quando quando si descrive la struttura terziaria, le caratteristiche strutturali della proteina (frammenti di amminoacidi, l'ordine della loro alternanza, legami idrogeno) non sono indicate e si presume che tutte le proteine contengano "dettagli " tratto da un set standard di venti aminoacidi (tabella 1). Il compito principale quando si raffigura una struttura terziaria è mostrare la disposizione spaziale e l'alternanza delle strutture secondarie.
Riso. 9 DIVERSE OPZIONI PER RAPPRESENTARE LA STRUTTURA DELLA PROTEINA CRUMBIN.
A – formula strutturale nell'immagine spaziale.
B – struttura sotto forma di modello tridimensionale.
B – struttura terziaria della molecola.
D – combinazione delle opzioni A e B.
D – immagine semplificata della struttura terziaria.
E – struttura terziaria con ponti disolfuro.
La più conveniente per la percezione è la struttura terziaria volumetrica (opzione B), liberata dai dettagli della formula strutturale.
Una molecola proteica con una struttura terziaria, di regola, assume una certa configurazione, che è formata da interazioni polari (elettrostatiche) e legami idrogeno. Di conseguenza, la molecola assume la forma di una palla compatta - proteine globulari (globuli, lat. palla), o filamentose - proteine fibrillari (fibra, lat. fibra).
Un esempio di struttura globulare è la proteina albumina; la classe dell'albumina comprende l'albume dell'uovo di gallina. La catena polimerica dell'albumina è composta principalmente da alanina, acido aspartico, glicina e cisteina, alternati in un certo ordine. La struttura terziaria contiene α-eliche collegate da singole catene (Fig. 10).
Riso. 10 STRUTTURA GLOBOLARE DELL'ALBUMINA
Un esempio di struttura fibrillare è la proteina fibroina. Contengono un gran numero di residui di glicina, alanina e serina (ogni secondo residuo aminoacidico è glicina); non sono presenti residui di cisteina contenenti gruppi solfidrico. La fibroina, il componente principale della seta naturale e delle tele dei ragni, contiene strutture β collegate da singole catene (Fig. 11).
Riso. 11 PROTEINA FIBRILARE FIBROINA
La possibilità di formare una struttura terziaria di un certo tipo è inerente alla struttura primaria della proteina, cioè determinato in anticipo dall'ordine di alternanza dei residui aminoacidici. Da alcuni insiemi di tali residui si formano prevalentemente α-eliche (ce ne sono molti di questi insiemi), un altro insieme porta alla comparsa di strutture β, le singole catene sono caratterizzate dalla loro composizione.
Alcune molecole proteiche, pur mantenendo la loro struttura terziaria, sono in grado di combinarsi in grandi aggregati sopramolecolari, mentre sono tenute insieme da interazioni polari e da legami idrogeno. Tali formazioni sono chiamate struttura quaternaria della proteina. Ad esempio, la proteina ferritina, che consiste principalmente di leucina, acido glutammico, acido aspartico e istidina (la ferricina contiene tutti i 20 residui aminoacidici in quantità variabili), forma una struttura terziaria di quattro eliche α disposte parallelamente. Quando le molecole vengono combinate in un unico insieme (Fig. 12), si forma una struttura quaternaria, che può includere fino a 24 molecole di ferritina.
Fig.12 FORMAZIONE DELLA STRUTTURA QUATERNARIA DELLA PROTEINA GLOBOLARE FERRITINA
Un altro esempio di formazioni supramolecolari è la struttura del collagene. È una proteina fibrillare, le cui catene sono costituite principalmente da glicina, alternata a prolina e lisina. La struttura contiene catene singole, triple eliche α, alternate a strutture β a forma di nastro disposte in fasci paralleli (Fig. 13).
Fig.13 STRUTTURA SUPRAMOLECOLARE DELLA PROTEINA FIBRILARE DEL COLLAGENE
Proprietà chimiche delle proteine.
Sotto l'azione di solventi organici, prodotti di scarto di alcuni batteri (fermentazione dell'acido lattico) o con l'aumento della temperatura, la distruzione delle strutture secondarie e terziarie avviene senza danneggiare la sua struttura primaria, per cui la proteina perde solubilità e perde attività biologica, questo processo è chiamato denaturazione, cioè la perdita di proprietà naturali, ad esempio la cagliatura del latte acido, l'albume coagulato di un uovo di gallina bollito. A temperature elevate, le proteine degli organismi viventi (in particolare i microrganismi) si denaturano rapidamente. Tali proteine non sono in grado di partecipare ai processi biologici, di conseguenza i microrganismi muoiono, quindi il latte bollito (o pastorizzato) può essere conservato più a lungo.
I legami peptidici H-N-C=O che formano la catena polimerica di una molecola proteica vengono idrolizzati in presenza di acidi o alcali, causando la rottura della catena polimerica, che alla fine può portare agli amminoacidi originali. I legami peptidici che fanno parte delle eliche α o delle strutture β sono più resistenti all'idrolisi e a vari influssi chimici (rispetto agli stessi legami nelle catene singole). Un disassemblaggio più delicato della molecola proteica negli amminoacidi che la compongono viene effettuato in un ambiente anidro utilizzando idrazina H 2 N–NH 2 , mentre tutti i frammenti di amminoacidi, tranne l'ultimo, formano le cosiddette idrazidi dell'acido carbossilico contenenti il frammento C(O)–HN–NH2 (Fig. 14).
Riso. 14. DIVISIONE POLIPEPTIDE
Tale analisi può fornire informazioni sulla composizione aminoacidica di una particolare proteina, ma è più importante conoscerne la sequenza nella molecola proteica. Uno dei metodi largamente utilizzati a questo scopo è l'azione del fenil isotiocianato (FITC) sulla catena polipeptidica, che in ambiente alcalino è attaccata al polipeptide (dall'estremità che contiene il gruppo amminico), e quando la reazione del l'ambiente diventa acido, si stacca dalla catena portando con sé un frammento di un amminoacido (Fig. 15).
Riso. 15 Scissione sequenziale del polipeptide
Per tale analisi sono state sviluppate molte tecniche speciali, comprese quelle che iniziano a “smontare” la molecola proteica nei suoi componenti costitutivi, a partire dall'estremità carbossilica.
I ponti disolfuro incrociati S-S (formati dall'interazione dei residui di cisteina, Fig. 2 e 9) vengono scissi, convertendoli in gruppi HS mediante l'azione di vari agenti riducenti. L'azione degli agenti ossidanti (ossigeno o acqua ossigenata) porta nuovamente alla formazione di ponti disolfuro (Fig. 16).
Riso. 16. FISSAZIONE DEI PONTI DISOLFURO
Per creare ulteriori legami incrociati nelle proteine, viene utilizzata la reattività dei gruppi amminici e carbossilici. I gruppi amminici che si trovano nella struttura laterale della catena sono più accessibili a varie interazioni: frammenti di lisina, asparagina, lisina, prolina (Tabella 1). Quando tali gruppi amminici interagiscono con la formaldeide, avviene un processo di condensazione e compaiono i ponti trasversali –NH–CH2–NH– (Fig. 17).
Riso. 17 CREAZIONE DI ULTERIORI PONTI CROCIATI TRA MOLECOLE PROTEICHE.
I gruppi carbossilici terminali della proteina sono in grado di reagire con composti complessi di alcuni metalli polivalenti (i composti del cromo sono più spesso utilizzati) e si verificano anche legami incrociati. Entrambi i processi sono utilizzati nella concia della pelle.
Il ruolo delle proteine nel corpo.
Il ruolo delle proteine nel corpo è vario.
Enzimi(fermentazione lat. – fermentazione), il loro altro nome è enzimi (en zumh greco. - nel lievito) sono proteine ad attività catalitica in grado di aumentare migliaia di volte la velocità dei processi biochimici; Sotto l'azione degli enzimi, i componenti costitutivi del cibo: proteine, grassi e carboidrati vengono scomposti in composti più semplici, dai quali vengono poi sintetizzate nuove macromolecole necessarie per un certo tipo di organismo. Gli enzimi prendono parte anche a molti processi di sintesi biochimica, ad esempio alla sintesi delle proteine (alcune proteine aiutano a sintetizzarne altre). Cm. ENZIMI
Gli enzimi non sono solo catalizzatori altamente efficienti, ma anche selettivi (dirigono la reazione rigorosamente in una determinata direzione). In loro presenza, la reazione procede con una resa quasi del 100% senza formazione di sottoprodotti e le condizioni sono miti: pressione atmosferica e temperatura normali di un organismo vivente. Per fare un confronto, la sintesi dell'ammoniaca da idrogeno e azoto in presenza di un catalizzatore - ferro attivato - viene effettuata a 400–500 ° C e una pressione di 30 MPa, la resa di ammoniaca è del 15–25% per ciclo. Gli enzimi sono considerati catalizzatori senza rivali.
La ricerca intensiva sugli enzimi è iniziata a metà del XIX secolo. Oggi sono stati studiati più di 2000 enzimi diversi, che rappresentano la classe di proteine più diversificata.
I nomi degli enzimi sono composti come segue: la desinenza -asi viene aggiunta al nome del reagente con cui interagisce l'enzima, o al nome della reazione catalizzata, ad esempio l'arginasi decompone l'arginina (Tabella 1), la decarbossilasi catalizza la decarbossilazione , cioè. rimozione di CO2 dal gruppo carbossilico:
– COOH → – CH + CO2
Spesso, per indicare con maggiore precisione il ruolo di un enzima, nel suo nome vengono indicati sia l'oggetto che il tipo di reazione, ad esempio l'alcol deidrogenasi, un enzima che effettua la deidrogenazione degli alcoli.
Per alcuni enzimi, scoperti molto tempo fa, è stato conservato il nome storico (senza la desinenza –aza), ad esempio pepsina (pepsis, greco. digestione) e la tripsina (thrypsis greco. liquefazione), questi enzimi scompongono le proteine.
Per la sistematizzazione, gli enzimi sono raggruppati in grandi classi, la classificazione si basa sul tipo di reazione, le classi sono denominate secondo il principio generale - il nome della reazione e il finale - aza. Alcune di queste classi sono elencate di seguito.
Ossidoreduttasi– enzimi che catalizzano le reazioni redox. Le deidrogenasi incluse in questa classe effettuano il trasferimento di protoni, ad esempio l'alcol deidrogenasi (ADH) ossida gli alcoli in aldeidi, la successiva ossidazione delle aldeidi in acidi carbossilici è catalizzata dalle aldeide deidrogenasi (ALDH). Entrambi i processi avvengono nel corpo durante la conversione dell'etanolo in acido acetico (Fig. 18).
Riso. 18 OSSIDAZIONE A DUE STADI DELL'ETANOLO all'acido acetico
Non è l'etanolo ad avere un effetto narcotico, ma il prodotto intermedio acetaldeide, quanto minore è l'attività dell'enzima ALDH, tanto più lento è il secondo stadio: l'ossidazione dell'acetaldeide in acido acetico e più lungo e forte l'effetto inebriante derivante dall'ingestione; etanolo. L'analisi ha mostrato che oltre l'80% dei rappresentanti della razza gialla hanno un'attività ALDH relativamente bassa e quindi hanno una tolleranza all'alcol notevolmente più grave. La ragione di questa ridotta attività congenita dell’ALDH è che alcuni dei residui di acido glutammico nella molecola “indebolita” dell’ALDH sono sostituiti da frammenti di lisina (Tabella 1).
Transferasi– enzimi che catalizzano il trasferimento di gruppi funzionali, ad esempio la transiminasi catalizza il movimento di un gruppo amminico.
Idrolasi– enzimi che catalizzano l’idrolisi. La tripsina e la pepsina precedentemente menzionate idrolizzano i legami peptidici e le lipasi scindono il legame estere nei grassi:
–RC(O)OR 1 +H 2 O → –RC(O)OH + HOR 1
Liasi– enzimi che catalizzano reazioni che non avvengono idroliticamente a seguito di tali reazioni, i legami C-C, C-O, C-N si rompono e si formano nuovi legami; A questa classe appartiene l'enzima decarbossilasi
Isomerasi– enzimi che catalizzano l'isomerizzazione, ad esempio la conversione dell'acido maleico in acido fumarico (Fig. 19), questo è un esempio di isomerizzazione cis - trans (vedi ISOMERIA).
Riso. 19. ISOMERIZZAZIONE DELL'ACIDO MALEICO in fumarico in presenza di un enzima.
Nel lavoro degli enzimi si osserva un principio generale secondo il quale esiste sempre una corrispondenza strutturale tra l'enzima e il reagente della reazione accelerata. Secondo l'espressione figurata di uno dei fondatori della dottrina degli enzimi, E. Fisher, il reagente si adatta all'enzima come una chiave per una serratura. A questo proposito, ciascun enzima catalizza una specifica reazione chimica o un gruppo di reazioni dello stesso tipo. A volte un enzima può agire su un singolo composto, ad esempio l'ureasi (uron greco. – urina) catalizza solo l’idrolisi dell’urea:
(H2N)2C = O + H2O = CO2 + 2NH3
La selettività più sottile è esibita dagli enzimi che distinguono tra antipodi otticamente attivi: isomeri destrorsi e mancini. La L-arginasi agisce solo sull'arginina levogira e non influenza l'isomero destrogiro. La L-lattato deidrogenasi agisce solo sugli esteri levogiri dell'acido lattico, i cosiddetti lattati (lactis lat. latte), mentre la D-lattato deidrogenasi scompone esclusivamente i D-lattati.
La maggior parte degli enzimi non agisce su uno, ma su un gruppo di composti correlati, ad esempio la tripsina “preferisce” scindere i legami peptidici formati da lisina e arginina (Tabella 1).
Le proprietà catalitiche di alcuni enzimi, come le idrolasi, sono determinate esclusivamente dalla struttura della molecola proteica stessa. Un'altra classe di enzimi: le ossidoreduttasi (ad esempio l'alcool deidrogenasi) possono essere attive solo in presenza di molecole non proteiche associate; loro - vitamine, ioni attivanti Mg, Ca, Zn, Mn e frammenti di acidi nucleici (Fig. 20).
Riso. 20 MOLECOLA DI ALCOL DEIDROGENASI
Le proteine di trasporto legano e trasportano varie molecole o ioni attraverso le membrane cellulari (sia all'interno che all'esterno della cellula), nonché da un organo all'altro.
Ad esempio, l’emoglobina lega l’ossigeno mentre il sangue passa attraverso i polmoni e lo consegna a vari tessuti del corpo, dove l’ossigeno viene rilasciato e quindi utilizzato per ossidare i componenti del cibo, questo processo funge da fonte di energia (a volte il termine “bruciare” di cibo nel corpo viene utilizzato).
Oltre alla parte proteica, l'emoglobina contiene un composto complesso di ferro con la molecola ciclica porfirina (porphyros greco. – viola), che provoca il colore rosso del sangue. È questo complesso (Fig. 21, a sinistra) che svolge il ruolo di trasportatore di ossigeno. Nell'emoglobina, il complesso del ferro porfirinico si trova all'interno della molecola proteica ed è mantenuto in posizione attraverso interazioni polari, nonché un legame di coordinazione con l'azoto nell'istidina (Tabella 1), che fa parte della proteina. La molecola di O2 trasportata dall'emoglobina è attaccata tramite un legame di coordinazione all'atomo di ferro sul lato opposto a quello a cui è attaccata l'istidina (Fig. 21, a destra).
Riso. 21 STRUTTURA DEL COMPLESSO DEL FERRO
La struttura del complesso è mostrata a destra sotto forma di modello tridimensionale. Il complesso è trattenuto nella molecola proteica da un legame di coordinazione (linea tratteggiata blu) tra l'atomo di Fe e l'atomo di N nell'istidina che fa parte della proteina. La molecola di O2 trasportata dall'emoglobina è attaccata in modo coordinato (linea tratteggiata rossa) all'atomo di Fe dal lato opposto del complesso planare.
L'emoglobina è una delle proteine più studiate; è costituita da a-eliche collegate da singole catene e contiene quattro complessi di ferro. Pertanto, l'emoglobina è come un voluminoso pacco per il trasporto di quattro molecole di ossigeno contemporaneamente. La forma dell'emoglobina corrisponde alle proteine globulari (Fig. 22).
Riso. 22 FORMA GLOBOLARE DELL'EMOGLOBINA
Il principale “vantaggio” dell’emoglobina è che l’aggiunta di ossigeno e la sua successiva eliminazione durante il trasferimento ai vari tessuti e organi avviene rapidamente. Il monossido di carbonio, CO (monossido di carbonio), si lega al Fe nell'emoglobina ancora più velocemente, ma, a differenza dell'O 2, forma un complesso difficile da distruggere. Di conseguenza, tale emoglobina non è in grado di legare l'O 2, il che porta (se vengono inalate grandi quantità di monossido di carbonio) alla morte del corpo per soffocamento.
La seconda funzione dell'emoglobina è il trasferimento della CO 2 espirata, ma nel processo di legame temporaneo dell'anidride carbonica non è l'atomo di ferro a partecipare, ma il gruppo N H 2 della proteina.
La “prestazione” delle proteine dipende dalla loro struttura, ad esempio, la sostituzione del singolo residuo aminoacidico dell’acido glutammico nella catena polipeptidica dell’emoglobina con un residuo di valina (una rara anomalia congenita) porta a una malattia chiamata anemia falciforme.
Esistono anche proteine di trasporto che possono legare grassi, glucosio e aminoacidi e trasportarli sia all'interno che all'esterno delle cellule.
Le proteine di trasporto di un tipo speciale non trasportano le sostanze stesse, ma svolgono le funzioni di "regolatore di trasporto", facendo passare determinate sostanze attraverso la membrana (la parete esterna della cellula). Tali proteine sono più spesso chiamate proteine di membrana. Hanno la forma di un cilindro cavo e, essendo incastonati nella parete della membrana, assicurano il movimento di alcune molecole o ioni polari nella cellula. Un esempio di proteina di membrana è la porina (Fig. 23).
Riso. 23 PROTEINE PORINE
Le proteine alimentari e di conservazione, come suggerisce il nome, servono come fonti di nutrimento interno, molto spesso per gli embrioni di piante e animali, nonché nelle prime fasi di sviluppo di organismi giovani. Tra le proteine alimentari figurano l'albumina (Fig. 10), il componente principale dell'albume, e la caseina, la principale proteina del latte. Sotto l'influenza dell'enzima pepsina, la caseina si coagula nello stomaco, garantendone la ritenzione nel tratto digestivo e un assorbimento efficace. La caseina contiene frammenti di tutti gli aminoacidi necessari all'organismo.
La ferritina (Fig. 12), presente nei tessuti animali, contiene ioni ferro.
Le proteine di stoccaggio includono anche la mioglobina, che è simile per composizione e struttura all'emoglobina. La mioglobina è concentrata principalmente nei muscoli, il suo ruolo principale è quello di immagazzinare l'ossigeno che le fornisce l'emoglobina. Si satura rapidamente di ossigeno (molto più velocemente dell'emoglobina) e quindi lo trasferisce gradualmente a vari tessuti.
Le proteine strutturali svolgono una funzione protettiva (pelle) o una funzione di supporto: tengono insieme il corpo in un unico insieme e gli conferiscono forza (cartilagine e tendini). Il loro componente principale è la proteina fibrillare del collagene (Fig. 11), la proteina più comune nel mondo animale nel corpo dei mammiferi, rappresentando quasi il 30% della massa totale delle proteine. Il collagene ha un'elevata resistenza alla trazione (la resistenza della pelle è nota), ma a causa del basso contenuto di legami incrociati nel collagene della pelle, le pelli animali nella loro forma grezza sono di scarsa utilità per la fabbricazione di vari prodotti. Per ridurre il rigonfiamento della pelle in acqua, il restringimento durante l'essiccazione, nonché per aumentare la resistenza allo stato acquoso e aumentare l'elasticità, vengono creati ulteriori collegamenti incrociati nel collagene (Fig. 15a), questo è il cosiddetto processo di concia della pelle .
Negli organismi viventi, le molecole di collagene che si formano durante la crescita e lo sviluppo dell'organismo non vengono rinnovate e non vengono sostituite da quelle di nuova sintesi. Con l'invecchiamento del corpo, aumenta il numero di legami incrociati nel collagene, il che porta ad una diminuzione della sua elasticità e, poiché non si verifica il rinnovamento, compaiono cambiamenti legati all'età: un aumento della fragilità della cartilagine e dei tendini e l'aspetto delle rughe sulla pelle.
I legamenti articolari contengono elastina, una proteina strutturale che si allunga facilmente in due dimensioni. La resilina proteica, che si trova nei punti cardine delle ali di alcuni insetti, ha la massima elasticità.
Formazioni cornee: capelli, unghie, piume, costituite principalmente da proteine cheratina (Fig. 24). La sua principale differenza è il notevole contenuto di residui di cisteina che formano ponti disolfuro, che conferiscono elevata elasticità (la capacità di ripristinare la forma originale dopo la deformazione) ai capelli e ai tessuti di lana.
Riso. 24. FRAMMENTO DI PROTEINA FIBRILARE CHERATINA
Per modificare in modo irreversibile la forma di un oggetto di cheratina, è necessario prima distruggere i ponti disolfuro utilizzando un agente riducente, dare una nuova forma, e poi creare nuovamente i ponti disolfuro utilizzando un agente ossidante (Fig. 16), questo è esattamente ciò che si fa, ad esempio, la permanente.
Con un aumento del contenuto di residui di cisteina nella cheratina e, di conseguenza, un aumento del numero di ponti disolfuro, la capacità di deformarsi scompare, ma appare un'elevata resistenza (le corna degli ungulati e i gusci delle tartarughe contengono fino al 18% di cisteina frammenti). Il corpo dei mammiferi contiene fino a 30 diversi tipi di cheratina.
La fibroina proteica fibrillare, correlata alla cheratina, secreta dai bruchi del baco da seta quando arricciano un bozzolo, così come dai ragni quando tessono una rete, contiene solo strutture β collegate da singole catene (Fig. 11). A differenza della cheratina, la fibroina non ha ponti disolfuro incrociati ed è molto resistente alla trazione (la resistenza per sezione trasversale unitaria di alcuni campioni di tessuto è superiore a quella dei cavi d'acciaio). A causa della mancanza di legami incrociati, la fibroina è anelastica (è noto che i tessuti di lana sono quasi resistenti alle sgualcature, mentre i tessuti di seta si sgualciscono facilmente).
Proteine regolatrici.
Le proteine regolatrici, più comunemente chiamate ormoni, sono coinvolte in vari processi fisiologici. Ad esempio, l'ormone insulina (Fig. 25) è costituito da due catene α collegate da ponti disolfuro. L’insulina regola i processi metabolici che coinvolgono il glucosio; la sua assenza porta al diabete.
Riso. 25 INSULINA PROTEICA
La ghiandola pituitaria del cervello sintetizza un ormone che regola la crescita del corpo. Esistono proteine regolatrici che controllano la biosintesi di vari enzimi nel corpo.
Le proteine contrattili e motorie danno al corpo la capacità di contrarsi, cambiare forma e muoversi, in particolare i muscoli. Il 40% della massa di tutte le proteine contenute nei muscoli è miosina (mys, myos, greco. – muscolo). La sua molecola contiene sia parti fibrillare che globulari (Fig. 26)
Riso. 26 MOLECOLA DI MIOSINA
Tali molecole si combinano in grandi aggregati contenenti 300-400 molecole.
Quando la concentrazione degli ioni calcio cambia nello spazio che circonda le fibre muscolari, si verifica un cambiamento reversibile nella conformazione delle molecole - un cambiamento nella forma della catena dovuto alla rotazione dei singoli frammenti attorno ai legami di valenza. Ciò porta alla contrazione e al rilassamento muscolare; il segnale per modificare la concentrazione degli ioni calcio proviene dalle terminazioni nervose delle fibre muscolari. La contrazione muscolare artificiale può essere causata dall'azione degli impulsi elettrici, portando ad un brusco cambiamento nella concentrazione degli ioni calcio. Su questo si basa la stimolazione del muscolo cardiaco per ripristinare la funzione cardiaca;
Le proteine protettive aiutano a proteggere il corpo dall'invasione di batteri, virus e dalla penetrazione di proteine estranee (il nome generale dei corpi estranei è antigeni). Il ruolo delle proteine protettive è svolto dalle immunoglobuline (un altro nome per loro è anticorpi); riconoscono gli antigeni che sono entrati nel corpo e si legano saldamente ad essi. Nel corpo dei mammiferi, compreso l'uomo, esistono cinque classi di immunoglobuline: M, G, A, D ed E, la loro struttura, come suggerisce il nome, è globulare, inoltre sono tutte costruite in modo simile. L'organizzazione molecolare degli anticorpi è mostrata di seguito utilizzando l'esempio dell'immunoglobulina di classe G (Fig. 27). La molecola contiene quattro catene polipeptidiche collegate da tre ponti disolfuro S-S (sono mostrati in Fig. 27 con legami di valenza ispessiti e simboli S grandi), inoltre, ciascuna catena polimerica contiene ponti disolfuro intracatena. Le due grandi catene polimeriche (in blu) contengono 400–600 residui di amminoacidi. Le altre due catene (in verde) sono lunghe quasi la metà e contengono circa 220 residui di amminoacidi. Tutte e quattro le catene sono disposte in modo tale che i gruppi terminali H 2 N siano diretti nella stessa direzione.
Riso. 27 RAPPRESENTAZIONE SCHEMATICA DELLA STRUTTURA DELLE IMMUNOGLOBULINE
Dopo che il corpo entra in contatto con una proteina estranea (antigene), le cellule del sistema immunitario iniziano a produrre immunoglobuline (anticorpi), che si accumulano nel siero del sangue. Nella prima fase, il lavoro principale è svolto dai tratti delle catene contenenti il terminale H 2 N (in Fig. 27, i tratti corrispondenti sono contrassegnati in azzurro e verde chiaro). Queste sono aree di cattura dell'antigene. Durante la sintesi dell'immunoglobulina, queste aree si formano in modo tale che la loro struttura e configurazione corrispondano al massimo alla struttura dell'antigene in avvicinamento (come la chiave di una serratura, come gli enzimi, ma i compiti in questo caso sono diversi). Pertanto, per ciascun antigene, come risposta immunitaria viene creato un anticorpo strettamente individuale. Nessuna proteina conosciuta può modificare la sua struttura in modo così “plastico” a seconda di fattori esterni, oltre alle immunoglobuline. Gli enzimi risolvono il problema della corrispondenza strutturale con il reagente in modo diverso - con l'aiuto di un gigantesco insieme di vari enzimi, tenendo conto di tutti i casi possibili, e le immunoglobuline ricostruiscono ogni volta di nuovo lo "strumento di lavoro". Inoltre, la regione cerniera dell'immunoglobulina (Fig. 27) fornisce alle due aree di cattura una certa mobilità indipendente, di conseguenza la molecola dell'immunoglobulina può “trovare” contemporaneamente i due siti più convenienti per la cattura nell'antigene per potersi catturare in modo sicuro; aggiustalo, questo ricorda le azioni di una creatura crostaceo.
Successivamente, viene attivata una catena di reazioni sequenziali del sistema immunitario del corpo, le immunoglobuline di altre classi vengono collegate, di conseguenza la proteina estranea viene disattivata e quindi l'antigene (microrganismo estraneo o tossina) viene distrutto e rimosso.
Dopo il contatto con l'antigene, la concentrazione massima di immunoglobulina viene raggiunta (a seconda della natura dell'antigene e delle caratteristiche individuali dell'organismo stesso) entro diverse ore (a volte diversi giorni). Il corpo conserva la memoria di tale contatto e, con un attacco ripetuto dello stesso antigene, le immunoglobuline si accumulano nel siero del sangue molto più velocemente e in quantità maggiori: si verifica l'immunità acquisita.
La suddetta classificazione delle proteine è alquanto arbitraria, ad esempio la proteina trombina, menzionata tra le proteine protettive, è essenzialmente un enzima che catalizza l'idrolisi dei legami peptidici, cioè appartiene alla classe delle proteasi.
Le proteine protettive spesso includono proteine del veleno di serpente e proteine tossiche di alcune piante, poiché il loro compito è proteggere il corpo dai danni.
Esistono proteine le cui funzioni sono così uniche che è difficile classificarle. Ad esempio, la proteina monellina, contenuta in una pianta africana, ha un sapore molto dolce ed è stata studiata come sostanza non tossica che potrebbe essere utilizzata al posto dello zucchero per prevenire l'obesità. Il plasma sanguigno di alcuni pesci antartici contiene proteine con proprietà antigelo, che impediscono il congelamento del sangue di questi pesci.
Sintesi proteica artificiale.
La condensazione degli amminoacidi che portano a una catena polipeptidica è un processo ben studiato. È possibile, ad esempio, effettuare la condensazione di un qualsiasi amminoacido o di una miscela di acidi e, di conseguenza, ottenere un polimero contenente unità identiche o unità diverse alternate in ordine casuale. Tali polimeri hanno poca somiglianza con i polipeptidi naturali e non hanno attività biologica. Il compito principale è combinare gli amminoacidi in un ordine rigorosamente definito e predeterminato al fine di riprodurre la sequenza dei residui di amminoacidi nelle proteine naturali. Lo scienziato americano Robert Merrifield ha proposto un metodo originale che ha permesso di risolvere questo problema. L'essenza del metodo è che il primo amminoacido è attaccato ad un gel polimerico insolubile, che contiene gruppi reattivi che possono combinarsi con i gruppi –COOH – dell'amminoacido. Il polistirene reticolato con gruppi clorometilici introdotti in esso è stato preso come substrato polimerico. Per evitare che l'amminoacido prelevato per la reazione reagisca con se stesso e per evitare che si unisca al gruppo H 2 N del substrato, il gruppo amminico di questo acido viene prima bloccato con un sostituente voluminoso [(C 4 H 9) 3 ] Gruppo 3 OS (O). Dopo che l'amminoacido si è attaccato al supporto polimerico, il gruppo bloccante viene rimosso e nella miscela di reazione viene introdotto un altro amminoacido, anch'esso provvisto di un gruppo H 2 N precedentemente bloccato. In un tale sistema è possibile solo l'interazione del gruppo H 2 N del primo amminoacido e del gruppo –COOH del secondo acido, che avviene in presenza di catalizzatori (sali di fosfonio). Successivamente, l'intero schema viene ripetuto, introducendo il terzo amminoacido (Fig. 28).
Riso. 28. SCHEMA PER LA SINTESI DI CATENE POLIPEPTIDICHE
Nell'ultima fase le catene polipeptidiche risultanti vengono separate dal supporto di polistirolo. Ora l'intero processo è automatizzato; esistono sintetizzatori automatici di peptidi che funzionano secondo lo schema descritto. Molti peptidi utilizzati in medicina e in agricoltura sono stati sintetizzati utilizzando questo metodo. È stato anche possibile ottenere analoghi migliorati di peptidi naturali con effetti selettivi e potenziati. Vengono sintetizzate alcune piccole proteine, come l’ormone insulina e alcuni enzimi.
Esistono anche metodi di sintesi proteica che copiano i processi naturali: sintetizzano frammenti di acidi nucleici configurati per produrre determinate proteine, quindi questi frammenti vengono incorporati in un organismo vivente (ad esempio in un batterio), dopodiché il corpo inizia a produrre le proteine proteina desiderata. In questo modo si ottengono quantità significative di proteine e peptidi difficili da raggiungere, nonché dei loro analoghi.
Le proteine come fonti alimentari.
Le proteine in un organismo vivente vengono costantemente scomposte nei loro amminoacidi originali (con la partecipazione indispensabile di enzimi), alcuni amminoacidi vengono trasformati in altri, quindi le proteine vengono nuovamente sintetizzate (anche con la partecipazione di enzimi), ad es. il corpo si rinnova costantemente. Alcune proteine (il collagene della pelle e dei capelli) non si rinnovano; il corpo le perde continuamente e ne sintetizza di nuove. Le proteine come fonti alimentari svolgono due funzioni principali: forniscono al corpo il materiale da costruzione per la sintesi di nuove molecole proteiche e, inoltre, forniscono al corpo energia (fonti di calorie).
I mammiferi carnivori (incluso l'uomo) ottengono le proteine necessarie da alimenti vegetali e animali. Nessuna delle proteine ottenute dal cibo viene incorporata nel corpo senza modifiche. Nel tratto digestivo, tutte le proteine assorbite vengono scomposte in aminoacidi e da essi vengono costruite le proteine necessarie per un particolare organismo, mentre dagli 8 acidi essenziali (Tabella 1), i restanti 12 possono essere sintetizzati nel corpo se vengono sintetizzati. non vengono forniti in quantità sufficiente con il cibo, ma gli acidi essenziali devono essere forniti tassativamente con il cibo. Il corpo riceve atomi di zolfo nella cisteina dall'aminoacido essenziale metionina. Alcune proteine si degradano, rilasciando l'energia necessaria per mantenere la vita, e l'azoto che contengono viene escreto dal corpo attraverso le urine. In genere il corpo umano perde 25–30 g di proteine al giorno, quindi gli alimenti proteici devono essere sempre presenti nella quantità richiesta. Il fabbisogno minimo giornaliero di proteine è di 37 g per gli uomini e di 29 g per le donne, ma l'apporto raccomandato è quasi il doppio. Quando si valutano i prodotti alimentari, è importante considerare la qualità delle proteine. In assenza o in un basso contenuto di aminoacidi essenziali, le proteine sono considerate di scarso valore, quindi tali proteine dovrebbero essere consumate in quantità maggiori. Pertanto, le proteine dei legumi contengono poca metionina, mentre le proteine del grano e del mais hanno un basso contenuto di lisina (entrambi aminoacidi essenziali). Le proteine animali (esclusi i collageni) sono classificate come prodotti alimentari completi. Un set completo di tutti gli acidi essenziali contiene caseina del latte, così come la ricotta e il formaggio che ne deriva, quindi una dieta vegetariana, se è molto rigorosa, ad es. “Senza latticini” richiede un maggiore consumo di legumi, noci e funghi per fornire all’organismo gli aminoacidi essenziali nelle quantità necessarie.
Gli aminoacidi e le proteine sintetici vengono utilizzati anche come prodotti alimentari, aggiungendoli ai mangimi che contengono aminoacidi essenziali in piccole quantità. Esistono batteri in grado di processare e assimilare gli idrocarburi del petrolio; in questo caso, per la completa sintesi proteica, necessitano di essere alimentati con composti contenenti azoto (ammoniaca o nitrati). La proteina così ottenuta viene utilizzata come mangime per bestiame e pollame. Una serie di enzimi - carboidrasi - viene spesso aggiunta al mangime degli animali domestici, che catalizzano l'idrolisi di componenti difficili da decomporre degli alimenti a base di carboidrati (le pareti cellulari dei raccolti di grano), a seguito dei quali gli alimenti vegetali vengono assorbiti più completamente.
Michail Levickij
PROTEINE (articolo 2)
(proteine), una classe di composti complessi contenenti azoto, i componenti più caratteristici e importanti (insieme agli acidi nucleici) della materia vivente. Le proteine svolgono numerose e varie funzioni. La maggior parte delle proteine sono enzimi che catalizzano le reazioni chimiche. Anche molti ormoni che regolano i processi fisiologici sono proteine. Le proteine strutturali come il collagene e la cheratina sono i componenti principali del tessuto osseo, dei capelli e delle unghie. Le proteine contrattili muscolari hanno la capacità di modificare la loro lunghezza utilizzando l'energia chimica per eseguire un lavoro meccanico. Le proteine includono anticorpi che legano e neutralizzano le sostanze tossiche. Alcune proteine che possono rispondere agli influssi esterni (luce, odore) fungono da recettori negli organi sensoriali che percepiscono l'irritazione. Molte proteine situate all'interno della cellula e sulla membrana cellulare svolgono funzioni regolatrici.
Nella prima metà del XIX secolo. molti chimici, e tra questi soprattutto J. von Liebig, giunsero gradualmente alla conclusione che le proteine rappresentano una classe speciale di composti azotati. Il nome “proteine” (dal greco protos - primo) fu proposto nel 1840 dal chimico olandese G. Mulder.
PROPRIETÀ FISICHE
Le proteine sono bianche allo stato solido, ma incolori in soluzione, a meno che non portino qualche tipo di gruppo cromoforo (colorato), come l'emoglobina. La solubilità in acqua varia notevolmente tra le diverse proteine. Cambia anche a seconda del pH e della concentrazione di sali nella soluzione, quindi è possibile selezionare le condizioni in cui una proteina precipiterà selettivamente in presenza di altre proteine. Questo metodo di "salatura" è ampiamente utilizzato per isolare e purificare le proteine. La proteina purificata spesso precipita fuori dalla soluzione sotto forma di cristalli.
Rispetto ad altri composti, il peso molecolare delle proteine è molto elevato: da diverse migliaia a molti milioni di dalton. Pertanto, durante l'ultracentrifugazione, le proteine vengono sedimentate e a velocità diverse. A causa della presenza di gruppi carichi positivamente e negativamente nelle molecole proteiche, si muovono a velocità diverse e in un campo elettrico. Questa è la base dell'elettroforesi, un metodo utilizzato per isolare singole proteine da miscele complesse. Le proteine vengono anche purificate mediante cromatografia.
PROPRIETÀ CHIMICHE
Struttura.
Le proteine sono polimeri, cioè molecole costruite come catene da unità monomeriche ripetitive, o subunità, il cui ruolo è svolto dagli alfa aminoacidi. Formula generale degli amminoacidi
dove R è un atomo di idrogeno o un gruppo organico.
Una molecola proteica (catena polipeptidica) può essere costituita solo da un numero relativamente piccolo di aminoacidi o da diverse migliaia di unità monomeriche. La combinazione di amminoacidi in una catena è possibile perché ciascuno di essi ha due diversi gruppi chimici: un gruppo amminico basico, NH2, e un gruppo carbossilico acido, COOH. Entrambi questi gruppi sono attaccati all'atomo di carbonio α. Il gruppo carbossilico di un amminoacido può formare un legame ammidico (peptidico) con il gruppo amminico di un altro amminoacido:
Dopo che due amminoacidi sono stati legati in questo modo, la catena può essere allungata aggiungendone un terzo al secondo amminoacido e così via. Come si può vedere dall'equazione precedente, quando si forma un legame peptidico, viene rilasciata una molecola d'acqua. In presenza di acidi, alcali o enzimi proteolitici la reazione procede in senso inverso: la catena polipeptidica viene scissa in amminoacidi con l'aggiunta di acqua. Questa reazione è chiamata idrolisi. L'idrolisi avviene spontaneamente ed è necessaria energia per collegare gli amminoacidi in una catena polipeptidica.
Un gruppo carbossilico e un gruppo ammidico (o un gruppo immidico simile nel caso dell'amminoacido prolina) sono presenti in tutti gli amminoacidi, ma le differenze tra gli amminoacidi sono determinate dalla natura del gruppo, o "catena laterale", che è indicato sopra con la lettera R. Il ruolo della catena laterale può essere svolto da un atomo di idrogeno, come l'amminoacido glicina, e da qualche gruppo voluminoso, come l'istidina e il triptofano. Alcune catene laterali sono chimicamente inerti, mentre altre sono marcatamente reattive.
Molte migliaia di amminoacidi diversi possono essere sintetizzati e molti amminoacidi diversi sono presenti in natura, ma solo 20 tipi di amminoacidi vengono utilizzati per la sintesi proteica: alanina, arginina, asparagina, acido aspartico, valina, istidina, glicina, glutammina, glutammico acido, isoleucina, leucina, lisina, metionina, prolina, serina, tirosina, treonina, triptofano, fenilalanina e cisteina (nelle proteine, la cisteina può essere presente come dimero - cistina). È vero che alcune proteine contengono altri amminoacidi oltre ai venti regolari, ma si formano come risultato della modifica di uno dei venti elencati dopo che è stato incluso nella proteina.
Attività ottica.
Tutti gli amminoacidi, ad eccezione della glicina, hanno quattro gruppi diversi attaccati all'atomo di carbonio α. Dal punto di vista della geometria, quattro diversi gruppi possono essere collegati in due modi, e di conseguenza ci sono due possibili configurazioni, o due isomeri, che stanno tra loro come un oggetto sta alla sua immagine speculare, cioè come la mano sinistra a destra. Una configurazione è detta mancinissima o mancina (L) e l'altra destrorsa o destrogira (D) perché i due isomeri differiscono nel senso di rotazione del piano della luce polarizzata. Solo gli L-amminoacidi si trovano nelle proteine (l'eccezione è la glicina; può essere trovata solo in una forma perché due dei suoi quattro gruppi sono uguali) e tutti sono otticamente attivi (perché esiste un solo isomero). Gli amminoacidi D sono rari in natura; si trovano in alcuni antibiotici e nella parete cellulare dei batteri.
Sequenza aminoacidica.
Gli amminoacidi in una catena polipeptidica non sono disposti in modo casuale, ma in un certo ordine fisso, ed è questo ordine che determina le funzioni e le proprietà della proteina. Variando l'ordine dei 20 tipi di aminoacidi, puoi creare un numero enorme di proteine diverse, così come puoi creare tanti testi diversi dalle lettere dell'alfabeto.
In passato, la determinazione della sequenza aminoacidica di una proteina richiedeva spesso diversi anni. La determinazione diretta è ancora un compito piuttosto laborioso, sebbene siano stati creati dispositivi che ne consentono l'esecuzione automatica. Di solito è più semplice determinare la sequenza nucleotidica del gene corrispondente e dedurre da essa la sequenza aminoacidica della proteina. Ad oggi sono già state determinate le sequenze aminoacidiche di molte centinaia di proteine. Le funzioni delle proteine decifrate sono generalmente note e questo aiuta a immaginare le possibili funzioni di proteine simili formate, ad esempio, nelle neoplasie maligne.
Proteine complesse.
Le proteine costituite solo da aminoacidi sono dette semplici. Spesso, però, alla catena polipeptidica è attaccato un atomo di metallo o qualche composto chimico che non sia un amminoacido. Tali proteine sono chiamate complesse. Un esempio è l'emoglobina: contiene porfirina di ferro, che ne determina il colore rosso e le permette di agire come trasportatore di ossigeno.
I nomi delle proteine più complesse indicano la natura dei gruppi attaccati: le glicoproteine contengono zuccheri, le lipoproteine contengono grassi. Se l'attività catalitica di un enzima dipende dal gruppo a cui è attaccato, allora si parla di gruppo prostetico. Spesso una vitamina svolge il ruolo di un gruppo prostetico o ne fa parte. La vitamina A, ad esempio, attaccata a una delle proteine della retina, ne determina la sensibilità alla luce.
Struttura terziaria.
Ciò che è importante non è tanto la sequenza aminoacidica della proteina stessa (la struttura primaria), ma il modo in cui questa è disposta nello spazio. Lungo l'intera lunghezza della catena polipeptidica, gli ioni idrogeno formano regolari legami idrogeno, che le conferiscono la forma di un'elica o di uno strato (struttura secondaria). Dalla combinazione di tali eliche e strati risulta una forma compatta dell'ordine successivo: la struttura terziaria della proteina. Intorno ai legami che tengono le unità monomeriche della catena sono possibili rotazioni con piccoli angoli. Pertanto, da un punto di vista puramente geometrico, il numero di possibili configurazioni per qualsiasi catena polipeptidica è infinitamente grande. In realtà, ogni proteina normalmente esiste in una sola configurazione, determinata dalla sua sequenza aminoacidica. Questa struttura non è rigida, sembra “respirare” - fluttua attorno ad una certa configurazione media. Il circuito è piegato in una configurazione in cui l'energia libera (la capacità di produrre lavoro) è minima, proprio come una molla rilasciata si comprime solo fino a uno stato corrispondente all'energia libera minima. Spesso una parte della catena è strettamente legata all’altra tramite legami disolfuro (–S–S–) tra due residui di cisteina. Questo è in parte il motivo per cui la cisteina svolge un ruolo particolarmente importante tra gli aminoacidi.
La complessità della struttura delle proteine è così grande che non è ancora possibile calcolare la struttura terziaria di una proteina, anche se se ne conosce la sequenza aminoacidica. Ma se è possibile ottenere cristalli proteici, la sua struttura terziaria può essere determinata mediante diffrazione dei raggi X.
Nelle proteine strutturali, contrattili e in alcune altre proteine, le catene sono allungate e diverse catene leggermente piegate che si trovano vicine formano fibrille; le fibrille, a loro volta, si piegano in formazioni più grandi: le fibre. Tuttavia, la maggior parte delle proteine in soluzione hanno forma globulare: le catene sono avvolte in un globulo, come il filo in un gomitolo. L'energia libera con questa configurazione è minima, poiché gli amminoacidi idrofobici (“idrorepellenti”) sono nascosti all'interno del globulo e gli amminoacidi idrofili (“attraenti l'acqua”) si trovano sulla sua superficie.
Molte proteine sono complessi di diverse catene polipeptidiche. Questa struttura è chiamata struttura quaternaria della proteina. La molecola dell'emoglobina, ad esempio, è composta da quattro subunità, ciascuna delle quali è una proteina globulare.
Le proteine strutturali, grazie alla loro configurazione lineare, formano fibre che hanno una resistenza alla trazione molto elevata, mentre la configurazione globulare permette alle proteine di entrare in interazioni specifiche con altri composti. Sulla superficie del globulo, quando le catene sono disposte correttamente, compaiono cavità di una certa forma in cui si trovano gruppi chimici reattivi. Se la proteina è un enzima, allora un'altra molecola, solitamente più piccola, di qualche sostanza entra in tale cavità, proprio come una chiave entra in una serratura; in questo caso la configurazione della nuvola elettronica della molecola cambia sotto l'influenza dei gruppi chimici presenti nella cavità, e questo la costringe a reagire in un certo modo. In questo modo l’enzima catalizza la reazione. Le molecole anticorpali hanno anche cavità nelle quali si legano diverse sostanze estranee e vengono così rese innocue. Il modello “serratura e chiave”, che spiega l’interazione delle proteine con altri composti, ci permette di comprendere la specificità di enzimi e anticorpi, ad es. la loro capacità di reagire solo con determinati composti.
Proteine in diversi tipi di organismi.
Anche le proteine che svolgono la stessa funzione in diverse specie di piante e animali e che quindi portano lo stesso nome hanno una configurazione simile. Tuttavia differiscono leggermente nella sequenza aminoacidica. Quando le specie divergono da un antenato comune, alcuni amminoacidi in determinate posizioni vengono sostituiti da mutazioni di altri. Le mutazioni dannose che causano malattie ereditarie vengono eliminate dalla selezione naturale, ma quelle benefiche o almeno neutre possono persistere. Più due specie sono vicine tra loro, minori sono le differenze riscontrate nelle loro proteine.
Alcune proteine cambiano in tempi relativamente brevi, altre sono molto conservate. Quest'ultimo include, ad esempio, il citocromo c, un enzima respiratorio presente nella maggior parte degli organismi viventi. Negli esseri umani e negli scimpanzé, le sequenze di aminoacidi sono identiche, ma nel citocromo c del grano solo il 38% degli aminoacidi era diverso. Anche confrontando uomo e batteri si può ancora notare la somiglianza dei citocromi c (le differenze riguardano il 65% degli aminoacidi), sebbene l'antenato comune di batteri e uomo vivesse sulla Terra circa due miliardi di anni fa. Al giorno d'oggi, il confronto delle sequenze di amminoacidi viene spesso utilizzato per costruire un albero filogenetico (genealogico), che riflette le relazioni evolutive tra diversi organismi.
Denaturazione.
La molecola proteica sintetizzata, ripiegandosi, acquisisce la sua configurazione caratteristica. Questa configurazione, tuttavia, può essere distrutta dal riscaldamento, dalla modifica del pH, dall'esposizione a solventi organici e anche semplicemente agitando la soluzione fino alla comparsa di bolle sulla sua superficie. Una proteina modificata in questo modo è detta denaturata; perde la sua attività biologica e solitamente diventa insolubile. Esempi ben noti di proteine denaturate sono le uova sode o la panna montata. Piccole proteine contenenti solo un centinaio di aminoacidi sono capaci di rinaturazione, cioè riacquisire la configurazione originale. Ma la maggior parte delle proteine si trasforma semplicemente in una massa di catene polipeptidiche aggrovigliate e non ripristina la loro configurazione precedente.
Una delle principali difficoltà nell'isolare le proteine attive è la loro estrema sensibilità alla denaturazione. Questa proprietà delle proteine trova utile applicazione nella conservazione degli alimenti: l'alta temperatura denatura irreversibilmente gli enzimi dei microrganismi e i microrganismi muoiono.
SINTESI PROTEICA
Per sintetizzare le proteine, un organismo vivente deve possedere un sistema di enzimi in grado di unire un amminoacido all'altro. È inoltre necessaria una fonte di informazioni per determinare quali aminoacidi dovrebbero essere combinati. Poiché nell’organismo esistono migliaia di tipi di proteine e ciascuna di esse è composta in media da diverse centinaia di aminoacidi, le informazioni richieste devono essere davvero enormi. Viene immagazzinato (in modo simile a come viene archiviata una registrazione su un nastro magnetico) nelle molecole di acido nucleico che compongono i geni.
Attivazione degli enzimi.
Una catena polipeptidica sintetizzata da aminoacidi non è sempre una proteina nella sua forma finale. Molti enzimi vengono sintetizzati prima come precursori inattivi e diventano attivi solo dopo che un altro enzima rimuove diversi amminoacidi a un'estremità della catena. Alcuni degli enzimi digestivi, come la tripsina, sono sintetizzati in questa forma inattiva; questi enzimi vengono attivati nel tratto digestivo in seguito alla rimozione del frammento terminale della catena. L'ormone insulina, la cui molecola nella sua forma attiva è costituita da due corte catene, è sintetizzato sotto forma di una catena, la cosiddetta. proinsulina. La parte centrale di questa catena viene quindi rimossa e i frammenti rimanenti si legano insieme per formare la molecola dell'ormone attivo. Le proteine complesse si formano solo dopo che uno specifico gruppo chimico si è attaccato alla proteina e questa aggiunta spesso richiede anche un enzima.
Circolazione metabolica.
Dopo aver somministrato all'animale amminoacidi marcati con isotopi radioattivi di carbonio, azoto o idrogeno, l'etichetta viene rapidamente incorporata nelle sue proteine. Se gli amminoacidi marcati smettono di entrare nel corpo, la quantità di marcatori nelle proteine inizia a diminuire. Questi esperimenti dimostrano che le proteine risultanti non vengono trattenute nel corpo fino alla fine della vita. Tutti loro, con poche eccezioni, sono in uno stato dinamico, vengono costantemente scomposti in amminoacidi e poi vengono nuovamente sintetizzati.
Alcune proteine si degradano quando le cellule muoiono e vengono distrutte. Ciò accade continuamente, ad esempio, con i globuli rossi e le cellule epiteliali che rivestono la superficie interna dell'intestino. Inoltre, la scomposizione e la risintesi delle proteine avviene anche nelle cellule viventi. Stranamente, si sa meno sulla scomposizione delle proteine che sulla loro sintesi. È chiaro, tuttavia, che la degradazione coinvolge enzimi proteolitici simili a quelli che scompongono le proteine in amminoacidi nel tratto digestivo.
L'emivita delle diverse proteine varia da alcune ore a molti mesi. L'unica eccezione sono le molecole di collagene. Una volta formati, rimangono stabili e non vengono rinnovati né sostituiti. Con il passare del tempo, però, alcune delle loro proprietà cambiano, in particolare l'elasticità, e poiché non si rinnovano, ciò si traduce in alcuni cambiamenti legati all'età, come la comparsa di rughe sulla pelle.
Proteine sintetiche.
I chimici hanno imparato da tempo a polimerizzare gli amminoacidi, ma gli amminoacidi sono combinati in modo disordinato, così che i prodotti di tale polimerizzazione somigliano poco a quelli naturali. È vero, è possibile combinare gli aminoacidi in un determinato ordine, il che rende possibile ottenere alcune proteine biologicamente attive, in particolare l'insulina. Il processo è piuttosto complicato e in questo modo è possibile ottenere solo quelle proteine le cui molecole contengono circa un centinaio di aminoacidi. È preferibile invece sintetizzare o isolare la sequenza nucleotidica di un gene corrispondente alla sequenza amminoacidica desiderata, e quindi introdurre questo gene in un batterio, che produrrà grandi quantità del prodotto desiderato mediante replicazione. Questo metodo, però, ha anche i suoi inconvenienti.
PROTEINE E NUTRIZIONE
Quando le proteine del corpo vengono scomposte in aminoacidi, questi possono essere nuovamente utilizzati per sintetizzare le proteine. Allo stesso tempo, gli aminoacidi stessi sono soggetti a degradazione, quindi non vengono completamente riutilizzati. È anche chiaro che durante la crescita, la gravidanza e la guarigione delle ferite, la sintesi proteica deve superare la degradazione. Il corpo perde continuamente alcune proteine; Queste sono le proteine dei capelli, delle unghie e dello strato superficiale della pelle. Pertanto, per sintetizzare le proteine, ogni organismo deve ricevere aminoacidi dal cibo.
Fonti di aminoacidi.
Le piante verdi sintetizzano tutti i 20 aminoacidi presenti nelle proteine da CO2, acqua e ammoniaca o nitrati. Molti batteri sono anche in grado di sintetizzare gli amminoacidi in presenza di zucchero (o qualche equivalente) e di azoto fisso, ma lo zucchero viene infine fornito dalle piante verdi. Gli animali hanno una capacità limitata di sintetizzare gli amminoacidi; ottengono aminoacidi mangiando piante verdi o altri animali. Nel tratto digestivo, le proteine assorbite vengono scomposte in amminoacidi, questi ultimi vengono assorbiti e da essi vengono costruite le proteine caratteristiche di un dato organismo. Nessuna delle proteine assorbite viene incorporata nelle strutture corporee come tali. L'unica eccezione è che in molti mammiferi alcuni anticorpi materni possono passare intatti attraverso la placenta nel flusso sanguigno fetale e attraverso il latte materno (soprattutto nei ruminanti) possono essere trasferiti al neonato immediatamente dopo la nascita.
Fabbisogno proteico.
È chiaro che per mantenere la vita, il corpo deve ricevere una certa quantità di proteine dal cibo. Tuttavia, l’entità di questa necessità dipende da una serie di fattori. Il corpo ha bisogno del cibo sia come fonte di energia (calorie) sia come materiale per costruire le sue strutture. Il bisogno di energia viene prima di tutto. Ciò significa che quando nella dieta ci sono pochi carboidrati e grassi, le proteine alimentari vengono utilizzate non per la sintesi delle proprie proteine, ma come fonte di calorie. Durante il digiuno prolungato, anche le proprie proteine vengono utilizzate per soddisfare il fabbisogno energetico. Se nella dieta ci sono abbastanza carboidrati, il consumo di proteine può essere ridotto.
Bilancio dell'azoto.
In media ca. Il 16% della massa totale delle proteine è costituito da azoto. Quando gli aminoacidi contenuti nelle proteine vengono scomposti, l'azoto in esse contenuto viene escreto dal corpo nelle urine e (in misura minore) nelle feci sotto forma di vari composti azotati. È quindi conveniente utilizzare un indicatore come il bilancio dell'azoto per valutare la qualità della nutrizione proteica, vale a dire la differenza (in grammi) tra la quantità di azoto che entra nel corpo e la quantità di azoto escreto ogni giorno. Con un'alimentazione normale in un adulto, queste quantità sono uguali. In un organismo in crescita, la quantità di azoto escreto è inferiore alla quantità ricevuta, vale a dire il saldo è positivo. Se nella dieta mancano proteine, il bilancio è negativo. Se la dieta contiene abbastanza calorie, ma non contiene proteine, il corpo risparmia proteine. Allo stesso tempo, il metabolismo delle proteine rallenta e l’utilizzo ripetuto degli aminoacidi nella sintesi proteica avviene con la massima efficienza possibile. Tuttavia, le perdite sono inevitabili e i composti azotati vengono comunque escreti nelle urine e in parte nelle feci. La quantità di azoto escreto dal corpo ogni giorno durante il digiuno proteico può servire come misura della carenza proteica giornaliera. È naturale supporre che introducendo nella dieta una quantità di proteine equivalente a questa carenza, si possa ripristinare l'equilibrio dell'azoto. Tuttavia, questo non è vero. Dopo aver ricevuto questa quantità di proteine, il corpo inizia a utilizzare gli aminoacidi in modo meno efficiente, quindi sono necessarie alcune proteine aggiuntive per ripristinare l'equilibrio dell'azoto.
Se la quantità di proteine nella dieta supera quanto necessario per mantenere l’equilibrio dell’azoto, non sembra esserci alcun danno. Gli aminoacidi in eccesso vengono semplicemente utilizzati come fonte di energia. Per fare un esempio particolarmente eclatante, gli eschimesi consumano pochi carboidrati e circa dieci volte la quantità di proteine necessaria per mantenere l’equilibrio dell’azoto. Nella maggior parte dei casi, tuttavia, l’utilizzo delle proteine come fonte di energia non è vantaggioso perché una data quantità di carboidrati può produrre molte più calorie rispetto alla stessa quantità di proteine. Nei paesi poveri, le persone ottengono le calorie dai carboidrati e consumano quantità minime di proteine.
Se il corpo riceve la quantità necessaria di calorie sotto forma di prodotti non proteici, la quantità minima di proteine per garantire il mantenimento dell'equilibrio dell'azoto è di ca. 30 g al giorno. Circa questa quantità di proteine è contenuta in quattro fette di pane o 0,5 litri di latte. Un numero leggermente maggiore è solitamente considerato ottimale; Si consigliano da 50 a 70 g.
Aminoacidi essenziali.
Finora le proteine erano considerate nel loro insieme. Nel frattempo, affinché avvenga la sintesi proteica, tutti gli aminoacidi necessari devono essere presenti nel corpo. Il corpo stesso dell’animale è in grado di sintetizzare alcuni aminoacidi. Si chiamano sostituibili perché non devono necessariamente essere presenti nella dieta: è importante solo che l'apporto complessivo di proteine come fonte di azoto sia sufficiente; poi, se mancano gli aminoacidi non essenziali, l'organismo può sintetizzarli a scapito di quelli presenti in eccesso. I restanti aminoacidi “essenziali” non possono essere sintetizzati e devono essere forniti all’organismo attraverso il cibo. Essenziale per l'uomo sono valina, leucina, isoleucina, treonina, metionina, fenilalanina, triptofano, istidina, lisina e arginina. (Sebbene l'arginina possa essere sintetizzata nell'organismo, è classificata come aminoacido essenziale perché non viene prodotta in quantità sufficienti nei neonati e nei bambini in crescita. D'altro canto, alcuni di questi aminoacidi derivanti dal cibo potrebbero diventare inutili per un adulto persona.)
Questo elenco di aminoacidi essenziali è più o meno lo stesso in altri vertebrati e persino negli insetti. Il valore nutrizionale delle proteine viene solitamente determinato somministrandole a ratti in crescita e monitorando l'aumento di peso degli animali.
Valore nutrizionale delle proteine.
Il valore nutrizionale di una proteina è determinato dall’amminoacido essenziale che è maggiormente carente. Illustriamolo con un esempio. Le proteine del nostro corpo contengono in media ca. 2% triptofano (in peso). Diciamo che la dieta comprende 10 g di proteine contenenti l'1% di triptofano e che contengono abbastanza altri aminoacidi essenziali. Nel nostro caso 10 g di questa proteina incompleta equivalgono sostanzialmente a 5 g di proteina completa; i restanti 5 g possono servire solo come fonte di energia. Si noti che poiché gli aminoacidi non sono praticamente immagazzinati nel corpo e affinché avvenga la sintesi proteica, tutti gli aminoacidi devono essere presenti contemporaneamente, l'effetto dell'assunzione di aminoacidi essenziali può essere rilevato solo se tutti loro entrare nel corpo contemporaneamente.
La composizione media della maggior parte delle proteine animali è vicina alla composizione media delle proteine del corpo umano, quindi è improbabile che si verifichi una carenza di aminoacidi se la nostra dieta è ricca di alimenti come carne, uova, latte e formaggio. Tuttavia, ci sono proteine, come la gelatina (un prodotto della denaturazione del collagene), che contengono pochissimi aminoacidi essenziali. Le proteine vegetali, pur essendo in questo senso migliori della gelatina, sono anche povere di aminoacidi essenziali; Sono particolarmente poveri di lisina e triptofano. Tuttavia, una dieta puramente vegetariana non può essere considerata affatto dannosa, a meno che non consumi una quantità leggermente maggiore di proteine vegetali, sufficienti a fornire all’organismo gli aminoacidi essenziali. Le piante contengono la maggior parte delle proteine nei loro semi, soprattutto nei semi di grano e vari legumi. Anche i giovani germogli, come gli asparagi, sono ricchi di proteine.
Proteine sintetiche nella dieta.
Aggiungendo piccole quantità di aminoacidi essenziali sintetici o proteine ricche di aminoacidi a proteine incomplete, come le proteine del mais, il valore nutrizionale di queste ultime può essere significativamente aumentato, ad es. aumentando così la quantità di proteine consumate. Un'altra possibilità è quella di far crescere batteri o lieviti su idrocarburi del petrolio con l'aggiunta di nitrati o ammoniaca come fonte di azoto. La proteina microbica ottenuta in questo modo può servire come mangime per pollame o bestiame, oppure può essere consumata direttamente dall'uomo. Il terzo metodo, ampiamente utilizzato, utilizza la fisiologia dei ruminanti. Nei ruminanti, nella parte iniziale dello stomaco, il cosiddetto. Il rumine è abitato da forme speciali di batteri e protozoi che convertono le proteine vegetali incomplete in proteine microbiche più complete e queste, a loro volta, dopo la digestione e l'assorbimento, si trasformano in proteine animali. L’urea, un composto sintetico contenente azoto economico, può essere aggiunta al mangime per il bestiame. I microrganismi che vivono nel rumine utilizzano l'azoto ureico per convertire i carboidrati (di cui ce n'è molto di più nel mangime) in proteine. Circa un terzo di tutto l'azoto presente nell'alimentazione del bestiame può presentarsi sotto forma di urea, il che significa essenzialmente, in una certa misura, la sintesi chimica delle proteine.
Le proteine sono composti organici complessi costituiti da aminoacidi. L'analisi chimica ha mostrato che le proteine sono costituite dai seguenti elementi:
Carbonio 50-55%
Idrogeno 6-7%
Ossigeno 21-23%
Azoto 15-17%
Zolfo 0,3-2,5%.
Nella composizione delle singole proteine sono stati trovati anche fosforo, iodio, ferro, rame e altre macro e microsostanze.
Il contenuto degli elementi chimici di base può variare nelle singole proteine, ad eccezione dell'azoto, la cui quantità media è caratterizzata dalla massima costanza ed è del 16%. A questo proposito esiste un metodo per determinare la quantità di proteine in base all'azoto in esse contenuto. Sapendo che 6,25 grammi di proteine contengono 1 grammo di azoto, puoi trovare la quantità di proteine moltiplicando la quantità di azoto trovata per un fattore di 6,25.
2. 4. Aminoacidi.
Aminoacidi – acidi carbossilici il cui atomo di idrogeno di carbonio alfa è sostituito da un gruppo amminico. Le proteine sono costituite da aminoacidi. Attualmente si conoscono più di 200 aminoacidi diversi. Ce ne sono circa 60 nel corpo umano e le proteine contengono solo 20 aminoacidi, che vengono chiamati naturale o proteico. 19 di questi sono aminoacidi alfa, il che significa che un gruppo amminico è attaccato all'atomo di carbonio alfa dell'acido carbossilico. La formula generale di questi amminoacidi è la seguente.
Soltanto l'aminoacido prolina non corrisponde a questa formula; è classificato come imminoacido.
I nomi chimici degli amminoacidi sono abbreviati per brevità, ad esempio acido glutammico GLU, serina SEP, ecc. Recentemente, per scrivere la struttura primaria delle proteine sono stati utilizzati solo simboli di una lettera.
Tutti gli amminoacidi hanno gruppi comuni: -CH2, -NH2, -COOH, conferiscono proprietà chimiche generali alle proteine e ai radicali, la cui natura chimica è diversa. Determinano le caratteristiche strutturali e funzionali degli aminoacidi.
La classificazione degli amminoacidi si basa sulle loro proprietà fisico-chimiche.
Secondo la struttura dei radicali:
Ciclico - FEN omociclico, TIR, TRI eterociclico, GIS.
Aciclico - monoamminomonocarbossilico GLY, ALA, SER, CIS, TRE, MET, VAL, LEI, ILEY, NLEY, monoaminodicarbonico ASP, GLU, diamminomonocarbonico LIZ, ARG.
Per formazione nel corpo:
Sostituibile: può essere sintetizzato nel corpo da sostanze di natura proteica e non proteica.
Essenziali - non possono essere sintetizzati dall'organismo, quindi devono essere forniti solo con il cibo - tutti gli aminoacidi ciclici, TRE, VAL, LEI, IL.
Significato biologico degli aminoacidi:
Fanno parte delle proteine del corpo umano.
Fanno parte dei peptidi del corpo umano.
Molte sostanze biologicamente attive a basso peso molecolare si formano nel corpo da aminoacidi: GABA, ammine biogene, ecc.
Alcuni ormoni nel corpo sono derivati degli aminoacidi (ormoni tiroidei, adrenalina).
Precursori delle basi azotate che costituiscono gli acidi nucleici.
Precursori delle porfirine utilizzati per la biosintesi dell'eme per l'emoglobina e la mioglobina.
Precursori di basi azotate facenti parte di lipidi complessi (colina, etanolamina).
Partecipa alla biosintesi dei mediatori nel sistema nervoso (acetilcolina, dopamina, serotonina, norepinefrina, ecc.).
Proprietà degli aminoacidi:
Ben solubile in acqua.
In una soluzione acquosa esistono come una miscela di equilibrio di uno ione bipolare, forme cationiche e anioniche della molecola.
L'equilibrio dipende dal pH dell'ambiente.
NH3-CH-COOH NH3-CH-COO NH2-CH-COO
R + OH R R + H
Forma cationica Ione bipolare Forma anionica
pH alcalino Ambiente acido
Presentano proprietà anfotere.
Possono svolgere il ruolo di un sistema tampone, perché può reagire come una base debole e un acido debole.
 Interpretazione del sogno: perché sogni un cane in un sogno?
Interpretazione del sogno: perché sogni un cane in un sogno? Progetto di fisica sulle nanotecnologie
Progetto di fisica sulle nanotecnologie Creare una favola. Cos'è una favola? A. Glushkova, p. Alexandrovka
Creare una favola. Cos'è una favola? A. Glushkova, p. Alexandrovka